Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Ассоциация и биоценоз - Сукцессионная система - Био-экология, или синэкология
Выше использованы два термина “ассоциация” и “биоценоз”, вероятно, разумно обсудить их соответствие. В начале главы уже было сказано о неопределённости терминов “сообщество”, “биоценоз”, “биогеоценоз”, “экосистема”. Сообщество и биоценоз в русскоязычной литературе обычно рассматривают как синонимы. Экосистему в русскоязычной литературе понимают более широко, этот термин применяют и при рассмотрении явно искусственных единиц, для которых термины “сообщество”, “биоценоз” не используют. Биогеоценоз — это биоценоз + биотоп, т. е. с точки зрения биогеографии — это синонимы, ибо занимают одну территорию. Однако существование биоценоза как объекта, не имеющего биотопа невозможно, поэтому термин биогеоценоз явно избыточен.
В экологии существует две группы методов выделения биоценозов: по биоте и по биотопу.
Выделение биоценозов по биотопу наиболее просто, особенно широко его используют при выделении биоценозов, имеющих резкие отличия биотопа от окружения, например биоценоз озера посреди леса или острова в море. Такие методы широко используют при выделении биоценозов, образованных неподвижными или малоподвижными организмами. Выделение биоценозов ручьёв, родников или жёстких грунтов — широко распространённая практика экологических исследований пресных вод. В морской гидробиологии биоценозы выделяют по субстратам до сих пор, хотя и реже, чем ранее (Зернов, 1913; Дерюгин, 1915). В наземной экологии выделение биоценозов по биотопу уже почти не используют. Хотя в принципе нельзя отрицать возможность определения сообществ по биотопу, подобная диагностика тем точнее, чем лучше известно, какие характеристики биотопа наиболее значимы для сообществ. При решении этого вопроса имеет смысл особое внимание уделять тем характеристикам биотопа, которые меняет сукцессия. Очевидно, на современном уровне знаний возможна только очень грубая диагностика, гораздо менее точная, чем по видовому составу.
По биоте биоценозы (экосистемы, сообщества, биогеоценозы) на практике выделяют как некий результат математического анализа совокупности отобранных проб (пробных площадок и т.п.). При этом в анализе может быть использован только видовой состав или учитывают и обилие каждого вида. Выделение биоценозов на основе анализа видового состава практикуют в геоботанике, где оно восходит к Braun-Blanquet (1964). При анализе выделяют группы сопряжённых видов, диагностическим для выделения считают произвольно взятое число сопряжённых видов, чаще всего не менее 10. Выделение биоценозов с учётом обилия видов проводят в гидробиологии (Броцкая, Зенкевич, 1939; Шорыгин, 1955 и др.) и в геоботанике (Методические подходы..., 2010). При использовании количественных показателей встаёт вопрос о том, как оценивать доминирование. Проще всего использовать биомассу и численность. Эти показатели почти безраздельно доминируют в наземной экологии, где сообщества выделяют по доминирующим видам растительности. Нередко биомассу оценивают не в единицах массы, а в других величинах, например в проективном покрытии. Однако все эти показатели имеют недостатки. При выделении доминантов по биомассе таковыми становятся крупные, но медленно растущие виды. При использовании численности (этот показатель используют преимущественно в гидробиологии) в том же сообществе доминантом могут стать мелкие виды, имеющие низкую биомассу, но высокую численность. В геоботанике эти искажения отчасти нивелируют отдельным анализом разных ярусов растительности. Поскольку сообщество — функционирующий объект, правильнее для выделения доминирующих видов применить не структурные показатели (численность, биомассу), а функциональные. В качестве такого показателя в последнее время широко используют поток энергии через популяции видов. В наземной экологии этот показатель пока применяют мало, в первую очередь из-за методических трудностей, обусловленных очень крупными размерами деревьев. В гидробиологии, особенно морской, этот показатель, напротив, используют очень широко. Поток энергии можно считать пропорциональным интенсивности метаболизма популяции (трат на обмен). Интенсивность метаболизма популяции рассчитывают для животных по формуле:
R = c ∙ N ∙ Wd = c ∙ N1-d ∙Bd,
где W — средний индивидуальный сырой вес особи, N — численность и B — биомасса на 1 м2, d — аллометрический показатель, c — специфичный для таксона коэффициент удельной интенсивности метаболизма.
Если резко доминирующего вида нет, то используют разные методы многомерного анализа. В геоботанике для этих целей разработан математический аппарат и соответствующее программное обеспечение (обзор см. Методические подходы..., 2010). В гидробиологии используют стандартные методы многомерного анализа.
Все эти методы выделения сообществ основаны на группировке данных, собранных на разных станциях (пробах). При этом динамические аспекты (сукцесионная динамика) не учитывают, о них за редчайшим исключением даже не упоминают. Не удивительно, что выделенные группировки иногда удаётся интерпретировать биологически, иногда — нет. Впрочем, к тому, чтобы выделяемые биоценозы имели биологический смысл, не всегда и стремятся. Выделение сообществ на основе их физиономического сходства, кроме игнорирования сукцессионной динамики имеет имеет и другой врождённый порок: физиономически сходные участки поверхности Земли, имеющие разную сукцессионную динамику, относят к одному сообществу, хотя такое сходство конвергентно.
Необходимо также отметить, что использование в качестве показателя роли вида в сообществе потока энергии, проходящего через его популяцию, основано на идее о существовании вида-доминанта-эдификатора. Эта идея, казалось бы многократно подтверждённая фактическими данными, не имеет теоретического обоснования.
Виды, определяющие структуру и функционирование данного сообщества (key-species), вовсе не обязаны доминировать в нём ни по численности, ни по биомассе, ни по любому другому аналогичному показателю. Более того, они вовсе могут не быть учтены при использовании стандартных методов выделения сообществ. Например, существенную роль в поддержании травянистых сообществ играют виды, потребляющие растения, однако стандартные геоботанические методы полностью игнорируют такие виды. Другой пример: структуру сообществ макробентоса в значительной степени определяют виды, потребляющие макробентос, часто это рыбы, которых стандартные орудия сбора (дночерпатель, трал) не облавливают. Очевидно, что в этих случаях, а, вероятно, и во многих других правильнее было бы выделять сообщества по таким видам, но универсальных методических приёмов этого не существует. Такие виды можно установить только после изучения структуры и функционирования конкретных сообществ. Это требует гораздо больше ресурсов (людских, временных и материальных), поэтому на современном этапе развития экологии такие работы практически никогда не проводят.
“Ассоциация объединяет участки растительного покрова с одними и теми же господствующими видами каждого яруса и одной и той же сукцессионной тенденцией... все её участки при отсутствии нарушений или существенных изменений внешней среды сменяются участками одной и той же другой ассоциации (в ходе демутации) или одного и того же демутационного комплекса (в экогенезе). Окончательное установление ассоциаций — последняя стадия исследования района, к которой следует переходить, лишь выяснив его сукцессионные ряды” (Разумовский, 1981: 84).
Отдельная проблема — изучение сукцессий. Непосредственное их наблюдение из-за их длительности обычно невозможно. Методы, применимые в геоботанике описаны С.М. Разумовским (2011), некоторые из них можно использовать и в анализе бентосных проб. Например, раковины мёртвых моллюсков и остатки панцирей морских ежей обычно не перемещаются на значительные расстояния и по их составу можно было бы делать предположения о предшествующих ассоциациях и сукцессионной динамике. Методы изучения сукцессий на жёстких грунтах моря описаны В.В. Ошурковым (2000).
Хотя разные геоботанические школы трактуют “ассоциацию” неодинаково, её можно определить как элементарную (т. е. не допускающую дальнейшего деления) (син)экологическую (сукцессионную) единицу. В таком значении этот термин использован и здесь. Виды, входящие в ассоциацию взаимно адаптированы. В природе это проявляется как увеличение вероятности совместного нахождения. Выше на модели клеточных автоматов было показано, что только этого достаточно для образования участков с однородным населением даже в абсолютно однородной среде. Отдельные участки ассоциаций называют парцеллами. Парцеллу можно определить, как элементарную территориальную единицу. Ассоциации сменяют друг друга в ходе сукцессии, обычно плавно.
Выделяемые сообщества могут совпадать, а могут и не совпадать с ассоциациями. Степень этого совпадения часто зависит от соотношения величины учётной площадки и парцеллы и от скорости сукцессии. Если площади, занятые отдельными парцеллами меньше учётной площадки, то при обработке собранного материала в одно сообщество неизбежно будут объединены разные ассоциации. Кроме того, в одной парцелле могут сосуществовать (и обычно сосуществуют) виды нескольких ассоциаций, сменяющих друг друга в ходе сукцессии. Такое сообщество нельзя считать элементарной единицей, поскольку оно состоит из различных ассоциаций. Поэтому, используя или встречая термины “сообщество”, “экосистема” и т. п., необходимо понимать, к чему они относятся: к эволюционно сложившейся группе видов или к мозаике ассоциаций, объединённых в сообщество иногда только в результате несовершенства использованных методик сбора и обработки проб. В море такие комплексные сообщества, по-видимому, свойственны в первую очередь жёстким грунтам с их высокой мозаичностью. Например, как мозаику следует рассматривать заросли водорослей-макрофитов на жёстких грунтах и расположенные между ними на рыхлых грунтах ассоциации.
Мозаики, образованные одними и теми же ассоциациями, могут выглядеть как совершенно разные сообщества и при использовании стандартных методов даже не возникнут сомнения в этом. Пример такой ситуации приводит С.М. Разумовский (Рис. 1.41). Под Звенигородом “в сплошном массиве ельника... на повышениях располагались парцеллы ельника, а в понижениях — парцеллы двух стадий последней и предпоследней евтрофной гидросерии, лишённые древостоя из-за блокирующего действия ели. После первой вырубки... на парцеллах последней стадии гидросерии возобновилась осина, а парцеллы климакса и предпоследней стадии гидросерии оказались заблокированными. После второй вырубки..., напротив, предпоследняя стадия приобрела свойственный ей древостой из серой ольхи, заблокировавший климакс и последнюю стадию гидросерии” (Разумовский, 1981: 125). В результате, хотя мозаика ассоциаций не изменилась, стандартные геоботанические методы зарегистрировали смену сообществ. Однако эта смена — артефакт изпользования учётной площадки существенно превышающей размер элементов мозаики.
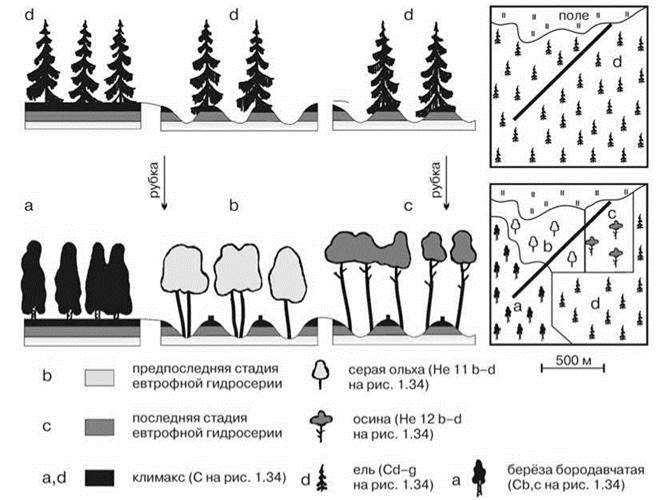
Рис. 1.41. Возникновение разных сообществ из одних и тех же парцелл. По Разумовскому (2011).
Сообщества обозначены разными буквенными индексами и силуэтами соответствующего цвета деревьев, парцеллы — различной заливкой на почвенных разрезах. Линией на правых рисунках показано положение разреза, изображённого на левых.
Хотя климакс — состояние ЦС, к которому она стремится, в реальных условиях из-за постоянно происходящих нарушений сукцессии никогда не приведут к климаксу на всей территории ЦС. Нормальным состоянием ЦС является мозаика ассоциаций разных сукцессионных рядов, находящихся на разных стадиях сукцессии. Более того, вообразим ситуацию, где климакс был бы достигнут на всей территории ЦС. Это означало бы не только установление климакса, но и исчезновение всех сериальных ассоциаций со всеми видами, входящими в их состав. И когда, рано или поздно, неизбежно произойдёт экзогенное нарушение климакса, восстанавливающие его сукцессии не смогут пойти, поскольку необходимые для этого виды вымерли. В результате климакс не сможет восстановиться. Поэтому установление климакса на всей территории ЦС неизбежно приведёт к её гибели.
Мозаичность ЦС создаётся не только экзогенными нарушениями, но и внутренними причинами. В ассоциациях с крупными и малоподвижными особями видов-эдификаторов, например, в лесных ассоциациях смерть каждого дерева приводит к существенному локальному изменению светового режима, часто нарушается и почвенный покров, появляется источник мёртвой органики и т. п. и т. д.** Поэтому реальный состав биоты на местности представляет собой динамически меняющуюся мозаику сукцессий восстановления. В геоботанике это нашло отражение в gap- концепции, которая, однако, оперирует значительно более крупными, объектами, состоящими из мозаики парцелл разных ассоциаций.