Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Возникновение ценотических (сукцессионных) систем - Сукцессионная система - Био-экология, или синэкология
Рассмотрим в эволюционном масштабе времени процесс возникновения ЦС на ранее безжизненной площади. Еще раз можно вспомнить случай Арала, показывающий, что “эволюционный масштаб времени” может быть всего лишь парой десятилетий. Разберем крайний случай: виды, заселяющие обозреваемую площадь, не имеют никаких адаптаций ни друг к другу, ни к специфическим условиям новой площади. Очевидно, что при первом же взаимодействии видов выявятся их макрониши (одни — автотрофы, другие — консументы первого порядка, третьи — хищники и т. п.). В пределах макрониш одни виды станут более обильными и обычными либо случайно, либо из-за того, что они окажутся конкурентоспособнее, чем другие. Такие виды будут в той или иной степени формировать окружающую среду, снижая её изменчивость (хотя бы лишь фактом своего наличия). Это приведет к возникновению ассоциаций видов. При достаточно высоком разнообразии условий на рассматриваемой площади в других её частях будут иные условия, где доступные ресурсы станут более подходящими для других видов. Они будут формировать свои ассоциации. Например, одни ассоциации станут возникать на рыхлых грунтах, другие — на скалах. Одни на переувлажнённых почвах, другие — на песках или сухих известняках. Т. е. первоначально бесструктурная в биотическом отношении площадь будет становиться всё более структурированной по мере формирования ассоциаций.
“По мере образования биоценозов с устойчивой структурой, растения более чётко распределяются по различным местообитаниям” (Толмачев, 1974: 212).
В силу чисто случайных причин (большая распространённость на данной площади одних условий и меньшая — других) одни из ассоциаций окажутся распространённее других. Их влияние на окружающие ассоциации будет более сильным в силу того, что более распространены, т. е. являются предсказуемым элементом окружения. С другой стороны, более распространённые ассоциации имеют больше возможностей для более тонкой и совершенной коадаптации входящих в него видов. В результате они будут становиться всё более распространёнными и оказывать всё более сильное влияние на остальные как всё более предсказуемый компонент среды. Процесс происходит, таким образом, с положительной обратной связью. В результате интеграция ЦС быстро растёт.
Ассоциации формирующейся ЦС, весьма вероятно, имеют несбалансированный обмен и их адаптации в первую очередь были направлены на выживание, а не на эффективность. Несбалансированность изменяет место, где находится сообщество (его биотоп): происходит накопление продуктов жизнедеятельности и/или перепотребление имевшихся изначально в данном месте ресурсов и т. д. Мидиевые банки заиляются из-за отложения фекалий и псевдофекалий. Озера мелеют из-за отложения ила и торфа. На бесплодных песках откладывается гумус. В результате ассоциация изменяет свой биотоп таким образом, что в данном месте более конкурентоспособной оказывается другая ассоциация, которая её сменяет. Эту смену ассоциаций под действием причин внутренних, появившихся в результате их функционирования, называют экогенетической сукцессией. Движущей силой экогенетической сукцессии, как и любой другой сукцессии, является нескомпенсированность воздействия популяций друг на друга. Необходимо отметить, что такие изменённые биотой биотопы могут существенно отличаться от имевшихся изначально на данной территории биотопов, т. е. функционирование биоты приводит к росту разнообразия условий. Это обеспечивает возможность роста биоразнообразия и тем больше, чем больше ассоциаций формируется.
Каждая ассоциация меняет свой биотоп одним единственным определённым образом (мидиевая банка заявляет грунт, а не откладывает гумус и не способствует зарастанию озера). Новому состоянию биотопа может более соответствовать другая ассоциация. Последовательность сменяющих друг друга во времени ассоциаций называют сукцессионный ряд (sere), а ассоциации, входящие в этот ряд, — сериальные (serai) ассоциации. Каждому достаточно отличному и распространённому исходному состоянию абиоты соответствует своя пионерная ассоциация и идущий от неё сукцессионный ряд. Сериальные ассоциации, подобно стадиям онтогенеза, неустойчивы во времени, поскольку являются лишь стадиями развития ЦС.
Примером сукцессионного ряда является зарастание некоторых низовых болот на территории Московского геоботанического района (Рис. 1.36).
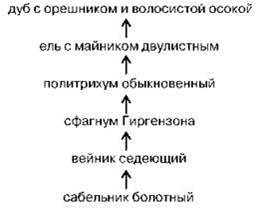
Рис. 1.36. Сукцессия зарастания низового болота в Московском ботанико-географическом районе. Ассоциации названы по доминанту. По С.М. Разумовскому (1981).
Ассоциации, входящие в ЦС, образуются не сразу и не одновременно. Анализ структуры изученных ЦС и их сукцессий показывает, что ранние стадии сукцессий часто образованы более древними эволюционно таксонами или же более древними ассоциациями. Так, в гидросериях (сукцессионных рядах наземных ЦС, образованных на переувлажненных субстратах) первые или одни из первых стадий образуют мхи. Ранние стадии восстановления современных рифов герматипных кораллов образованы водорослями — бурыми и красными известковыми. Развитие миоценовых коралловых рифов северо-восточной Атлантики, описанное в данной главе ниже, начинается с красных известковых водорослей. Палеонтологические данные показывают, что при формировании сообществ современных мелководных коралловых рифов геологически им предшествовали рифы красных известковых водорослей: хотя некоторые рода образующих их кораллов возникли в мелу, сами сообщества современных коралловых рифов возникли только в плиоцене—миоцене. Эта же закономерность повторялась всегда при неоднократном возникновении сообществ мелководных рифов в палеозое и мезозое (подробнее см. Жирков, 2010). Возможно, мы имеем дело с неким аналогом известного принципа “онтогенез повторяет филогенез”.
Сукцессионные ряды формируются только для достаточно распространённых стартовых условий абиоты. Редко встречающиеся сочетания факторов не имеют своих сукцессионных рядов, и такие местообитания осваивают ассоциации, сформировавшиеся в биотопах со сходными условиями. Если в ареале ЦС возникает достаточно обширная площадь с условиями ранее не встречавшимися или редкими, то вполне может возникнуть новый сукцессионный ряд или модифицироваться существующий. Иными словами, несмотря на существование жёсткой сукцессионной последовательности, ЦС не следует понимать, как окостеневшую структуру, она способна меняться вслед за изменением условий её существования.
Выше рассмотрена идеализированная ситуация, в которой ЦС формируется из видов, не имеющих никаких адаптаций ни друг к другу, ни к специфическим условиям новой площади и в отсутствии преемственности с ЦС, существовавшей в этом месте ранее. В действительности такая ситуация невероятна. В формирующейся биоте обычно существуют виды, преадаптированные к биотопам новой площади, они выступают “ядрами конденсации” образующихся сообществ. Ещё эффективнее роль таких ядер будут выполнять группы коадаптированных видов. Весьма вероятно, что эффективность роли ядер растёт нелинейно с увеличением числа входящих в них видов. Поскольку, обычно одна ЦС образуется из другой, число таких ядер во вновь формирующихся ЦС должно быть велико.
Основное отличие формирования новой ЦС на основе уже существовавших (одной или нескольких) — наличие большого числа уже сложившихся и коадаптированных комплексов видов. Часто в состав формирующейся ЦС могут даже входить в почти неизменном виде ассоциации материнских ЦС. Эти “ядра конденсации” должны существенно ускорять формирование новых ЦС. В принципе, новая ЦС может целиком состоять из “старых” видов, но обычно часть видов специфична для неё, ей присуща та или иная степень эндемизма, впрочем, виды обычно эндемичны не отдельной ЦС, а группе близких.
В ходе сукцессии несбалансированность обмена ассоциаций уменьшается. Это связано с тем, что сменяющие друг друга в ходе сукцессии ассоциации занимают одно и тоже место в пространстве, а количество поступающей на данное место энергии (света или детрита) постоянно. Всякая несбалансированность — это потери. Ассоциация, меньше теряющая энергию, оказывается в выигрыше. Также в результате сукцессии биотоп конкретной ассоциации видоизменяется таким образом, что отличия его от других уменьшаются. В результате сукцессионные ряды, начавшиеся в разных условиях с различных пионерных ассоциаций, сливаются друг с другом, постепенно приходя к ассоциации, которая в данных абиотических условиях (климат, глубина, обеспеченность пищей, освещённость и т. д. и т. п.) практически сбалансирована. Такую ассоциацию называют климаксной ассоциацией, или просто климаксом. В климаксе воздействие каждой популяции компенсирует воздействие остальных популяций. Климаксная ассоциация находится в состоянии динамического равновесия со своим биотопом. В результате изменение биотопа если и происходит, то несущественно с точки зрения самой климаксной ассоциации. Например, климаксовый коралловый риф может сколь угодно расти в размерах, но как был климаксом, так климаксом и останется. Единственный признак климакса, по которому его можно отличить от других ассоциаций — отсутствие у него внутренних причин для изменения. Итак, ЦС — это совокупность сукцессионных рядов (сериальных ассоциаций и климакса). Эта совокупность уникальна для каждой ЦС, хотя близкие ЦС могут иметь сходство.
В принципе нет ничего невероятного в том, что на территории одной сукцессионной системы сформируется сколько угодно климаксов. Весьма возможно, что при создании ЦС и начинает формироваться несколько климаксов. Несколько климаксов сосуществуют и при превращении субклимакса в климакс, и при удлинении сукцессионных рядов (см. ниже). Однако, поскольку каждый климакс и ведущие к нему сукцессионные ряды занимают определённую площадь, то между разными климаксами будет происходить конкурентная борьба за занимаемую площадь. Поэтому, для сосуществования нескольких климаксов в одной ЦС требуется или различие в требовании к биотопу или в ходе сукцессий. Примером первого варианта могут служить климаксы в бентали на рыхлых и жестких грунтах (см. Главу 5). Второго — безлесный и лесной климаксы (см. Главу 3).
При нарушении ассоциации внешними факторами возможны два варианта. Если вместе с ней нарушается и её биотоп, то сукцессия начинается с ассоциации, соответствующей новому состоянию биотопа.
Если нарушение затрагивает только ассоциацию, оставляя её биотоп нетронутым, происходит демутационная сукцессия — сравнительно быстрая последовательная смена нескольких недолговечных ассоциаций. Каждой достаточно продвинутой коренной ассоциации экогенетической сукцессии может соответствовать один или несколько демутационных сукцессионных рядов, состоящих из производных ассоциаций. Совокупность демутационных сукцессионных рядов и коренной ассоциации называют демутационный комплекс. Таким образом, экогенетическая сукцессия в действительности состоит не из последовательности ассоциаций, а последовательностей демутационных комплексов. Каждый демутационных комплекс, особенно в начале сукцессии, может состоять лишь из одной ассоциации. В качестве примера на Рис. 1.37 приведены демутационные сукцессии климакса Московского ботанико-географического района. Обратите внимание на то, что имеются две начальные стадии сукцессионных рядов. Один сукцессионный ряд начинается в том случае, если почвенный покров не поврежден (после верховых пожаров, вырубок, ветровалов и т. п.). Другой сукцессионный ряд начинается после повреждения почвенного покрова (после низовых пожаров, хищнических рубок с использованием тяжёлой техники и трелёвки бревен, в процессе которых почву часто сдирают до материнской породы). После прохождения нескольких стадий эти два сукцессионных ряда сливаются в один.
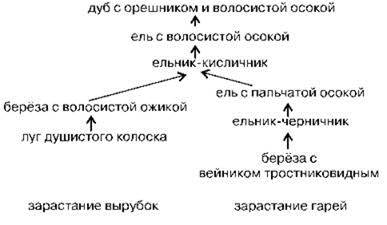
Рис. 1.37. Демутационные сукцессии климакса в Московском ботанико-географическом районе. По С.М. Разумовскому (1981).
Ассоциация демутационного комплекса, “которая занимает местообитание при отсутствии нарушений, называется коренной ассоциацией, а остальные, служащие стадиями ее демутационных смен, — производными ассоциациями” (Разумовский, 1981: 12).
Коренная ассоциация одинаково хорошо возобновляется как на собственном фоне, так и на фоне одной или нескольких производных ассоциаций. Ассоциации же демутационного ряда не способны возобновляться на собственном фоне: после естественного от возраста умирания их видов-эдификаторов одна ассоциация сменяет другую, а потому их существование ограничено продолжительностью жизни одного поколения видов-эдификаторов. Например, в демутационных ассоциациях с древесным ярусом отсутствует подрост соответствующих видов деревьев. Данное явление показано для всех изученных в этом отношении демутационных ассоциаций (Разумовский, 1981, 1999, 2011).
Широко известен феномен тропических лесов, в значительной степени обуславливающий их высокую мозаичность: под деревьями обычно отсутствует подрост того же вида. Для его объяснения было предложено несколько теорий, но ни одна из них не объясняет удовлетворительно все факты. Вполне вероятно, что тут имеют место демутационные сукцессии.
Ход сукцессий одновременно позволяет естественным образом разделить биоценоз и его биотоп. Под биоценозом следует понимать не только саму биоту, но и сумму всех отклоняющих воздействий, оказываемых ею на абиоту и бесследно исчезающих после устранения биоты. Например, малая скорость ветра в лесу или пониженная скорость течения в зарослях макрофитов является свойствами сообществ, а не их биотопов, поскольку обусловлены исключительно действием деревьев или макрофитов и исчезают, если убрать деревья или макрофиты.
В сообществах наземных растений сукцессии были детально рассмотрены С.М. Разумовским (1981). Он показал, что сукцессионные ряды различаются в зависимости от начальных условий, но все сукцессии приводят к климаксу, обычно одному в ЦС. Быстрее всего проходит демутационная сукцессия, но и её продолжительность не может быть меньше суммарной продолжительности жизни эдификаторов каждой стадии. Скорость прохождения сукцессий зависит как от видов, входящих в состав ассоциаций, так и от условий абиотической среды. Время существования различных ассоциаций может сильно различаться. Поскольку на суше эдификаторы всех сериальных ассоциаций, ведущих к лесному климаксу, за исключением нескольких пионерных, — деревья, то очевидно, что минимальное время даже демутационной сукцессии порядка 1000 лет. Время прохождения экогенетических сукцессий вероятно гораздо больше (тысячелетия, часто много тысячелетий), поскольку при их прохождении происходит не только смена доминантов, но и видоизменение биотопа. Как пример приведена сукцессионная система Московского геоботанического района (Рис. 1.38). В этой системе первые стадии зарастания песков (моховая и травяная) в сумме требуют полвека, а следующая стадия — сосняк с брусникой и ландышем — не менее 600 лет. Полное время прохождения сукцессии от пионерной стадии до климакса в Подмосковье составляет сотни, а в некоторых сукцессионных рядах, например, превращение озера или болота в лес, и тысячи лет. Очевидно, что время, прошедшее после разрушения человеком сообществ тундростепи (см. Главу 3), во многих случаях явно недостаточно для завершения экогенетических сукцессий. Интересующиеся детальным разбором приведённой схемы и аналогичных схем других районов могут обратиться к уже цитированным “Трудам...” С.М. Разумовского (2011).
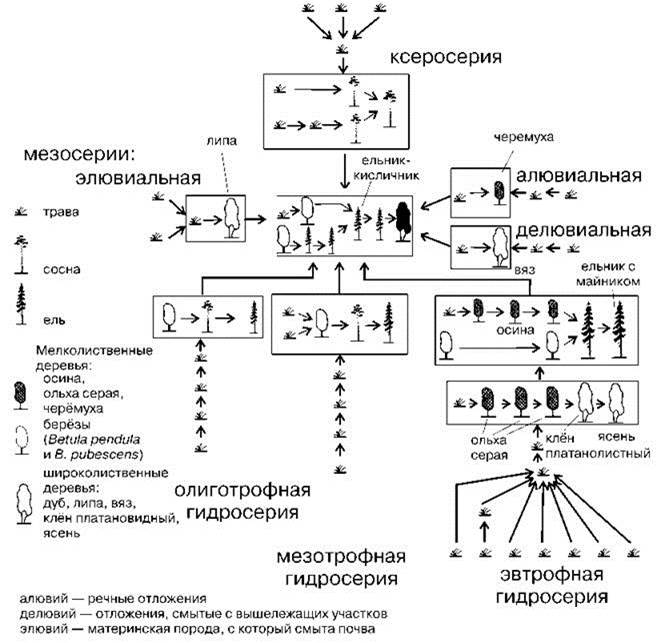
Рис. 1.38. Сукцессионная система Московского ботанико-географического района. По С.М. Разумовскому (1981).
Система имеет 7 основных экогенетических сукцессионных рядов, которые образованы 69 ассоциациями (показаны значками), 37 экогенетическими стадиями, из них 9 — демутационные комплексы (обведены рамками, в серой рамке — демутационный комплекс климакса). В случае двух демутационных рядов в одном демутационном комплексе верхний из них — вырубки, ветровалы, верховые пожары и другие воздействия, уничтожающие верхний ярус сообщества, но не затрагивающие подстилку, а нижний — с нарушением подстилки. Он часто возникает после низовых пожаров, но может быть вызван и любым другим воздействием, уничтожающим не только древостой, но и подстилку (трелевка с кронами, сбор подстилки населением, выпас в лесах, на крутых склонах и т. п.). Гидросерии — это зарастание верховых или низовых болот. Ксеросерия — зарастание песков. Аллювиальная мезосерия — зарастание речных отложений, делювиальная — отложений, смытых со склона, элювиальная — материнской породы, которая осталась вверху склона, после того, как с неё смыло почву.
Эвтрофная гидросерия и демутации климакса рассмотрены также на Рис. 1.36 и Рис. 1.37.
См. также Рис. 2.27 и 4.8.
Сходную картину показывает и изучение сукцессий в бентосе. Экспериментами по обрастанию пластин В.В. Ошурковым (2000) установлено, что на пластинах довольно быстро формируется ассоциация двустворчатого моллюска мидии Mylilus edulis. Если субстрат помещают в воду до или во время массового оседания спата (личинок мидий), на нём сразу формируется ассоциация мидий. Если позже, когда в планктоне нет личинок мидий, то образуются группировки гидроидов, усоногих раков или однолетних макрофитов (в зависимости от времени помещения субстрата, т. е. наличия в планктоне личинок соответствующих видов, и глубины), которые сменяет сообщество мидий, как только личинки мидий начинают оседать из планктона. Это именно группировки, а не ассоциации, поскольку их наличие для начала сукцессии необязательно. Исход сукцессии не зависит от того, были ли они на начальной стадии или нет. Стадия мидиевого сообщества длится много лет и заканчивается постепенным отмиранием мидий, достигших предельного возраста (в Белом море — 16 и более лет). На смену ассоциации мидий на глубинах 0—1 м приходят ассоциации фукоидов, 1—4 м — бурой водоросли Laminariasaccharina, через 2—3 года начинают появляться проростки L. digitata, но содоминантом ассоциации она становится лишь через 10—15 лет, когда большая часть её талломов достигнет размеров, сопоставимых с таковыми L. saccharina. Климаксной стадией сукцессии можно считать бидоминантную ассоциацию L. saccharina — L. digitata. Трёхъярусная структура фитоценоза и характерный флористический состав сопутствующих видов формируются в течение 3—4 лет (Михайлова, 2006). Глубже 4 м климаксной является ассоциация асцидии Styela rustica (Рис. 1.39).
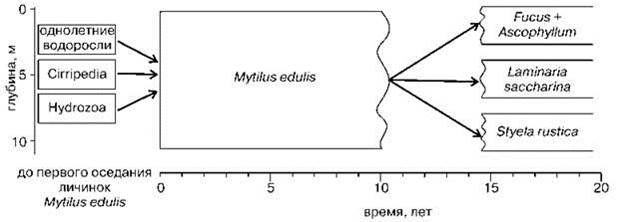
Рис. 1.39. Сукцессии эпифауны верхней сублиторали Белого моря. По данным В.В. Ошуркова (2000).
Аналогичные данные были получены и при изучении обрастаний прикамчатских вод Тихого океана (Ошурков, 2000). Сравнение данных по обрастаниям с наблюдениями в природе позволило В.В. Ошуркову (2000) сделать вывод о принципиальном сходстве сукцессий обрастания и сукцессий эпифаунных сообществ твёрдых грунтов. Везде в высокобореальных и субарктических водах экогенетические сукцессии приводят к смене ассоциации мидий ассоциациями бурых водорослей. В свою очередь, морские ежи Strongylocentrotus эффективно снижают плотность бурых водорослей, временами они способны полностью их уничтожить. Это приводит к замене ассоциаций бурых водорослей ассоциациями инкрустирующих багрянок. Для развития полноценных ассоциаций инкрустирующих багрянок, судя по зарастанию лав от извержений известного возраста, требуется не менее нескольких десятков лет (Ошурков, 2000). Пресс хищников способен эффективно снизить плотность популяций морских ежей и эндогенно стабилизировать ассоциации бурых водорослей. Каланы способны регулировать плотность ежей на глубинах до 15—20 м. Аналогичные данные получены и для калифорнийских келпов (Halpernet al., 2006), где ассоциацию бурых водорослей стабилизируют лангуст Panulirus interruptus и брюхоногий моллюск Kelletia kelletii (Buccinidae). Возможно, впрочем, что такая стабилизация не абсолютна: густые заросли бурых водорослей существенно снижают гидродинамику, что приводит к росту площадей дна, покрытых илом и песком. Это в свою очередь препятствует эффективному возобновлению зарослей, поскольку для прикрепления бурым водорослям необходимы твёрдые грунты (валуны, скалы). После исчезновения зарослей гидродинамика усиливается, и рыхлые грунты сменяют твёрдые. Другой возможный ход сукцессии — смена бурых водорослей зарослями морских трав.
Описана экогенетическая сукцессия миоценовых рифов северо-восточной Атлантики (Рис. 1.40). Ранние стадии аналогичной сукцессии описаны в прибрежных водах Сирии на глубинах 25—90 м. На илистых грунтах длинные, обильно ветвящиеся, стелющиеся по поверхности субстрата ризомы и многочисленные ризоиды зелёных сифоновых водорослей Caulerpa scalpelliformis и Udotea petiolata образуют плёнку. Поверхность водорослей покрыта клейкой полисахаридной слизью. На неё налипают членики мёртвых кораллиновых водорослей, кусочки раковин и панцирей бентосных животных, мелкие камешки и песчинки. На этих обломах поселяются корковые известковые (Lithophyllumracemus, Lithothamnion sp., Pseudolitophyllum expansum, Neogoniolithon sp.) и мягкие водоросли (бурая Zanardinia prototypus, красные Hildenbrandtia prototypus, Peyssonneliasp.), кустистая Amphiroa rigida, а также прикреплённые животные (мшанки, губки, гидроиды, асцидии, полихеты). Так формируется второй слой — прочная корка толщиной до 10 см. В результате формируются мостовые площадью в десятки квадратных километров (Maximova, Saker Fayes, 1999). В миоценовых рифах “на мобильном субстрате мелководий (до 35—40 м глубины) ядра из детрита обрастали Lithophyllum, создавая мелкие родолиты, на которых затем селились ветвистые Lithothamnion, а крупные родолиты могли покрываться листоватыми корками Mesophyllum” (Невесская, 1999: 292). Отдельные участки этой ассоциации могли разрастаться, образуя родолитовые мостовые. На таком известковом основании развивались рифы герматипных кораллов: сначала “возникали изолированные рифы, построенные Porites, на которых местами нарастали колонии Platygyra, а затем вся постройка покрывалась колониями Tarbellastraea. Кораллиновые водоросли, среди которых преобладали ветвистые Lithophyllum, росли вместе с кораллами только на первых стадиях формирования рифов, а во время господства Tarbellastraea исчезали. Эта сукцессия: кораллиновые водоросли Porites (Platygyra) Tarbellastraea могла повторяться несколько раз... Tarbellastraea могли селиться только на субстрате, уже занятом Porites. За гибелью Tarbellastraea из-за заноса осадком могло начаться вновь заселение субстрата представителями Porites. Каждый цикл длился несколько сотен лет” (Невесская, 1999: 292—293). Какие внешние воздействия приводили к заносу осадками рифа Tarbellastraea, возвращая сукцессию на уже прошедшие стадии, теперь уже вряд ли удастся установить.
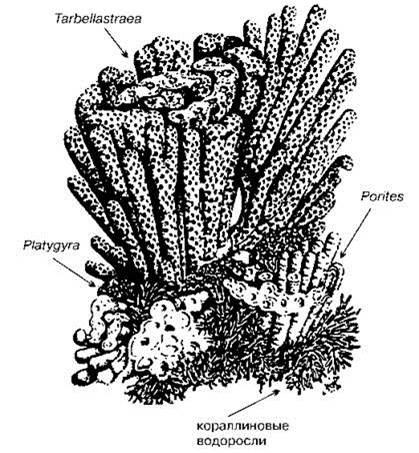
Рис. 1.40. Схема стадий роста рифа в тортонском море миоцена юго-восточной Испании. По Braga, Martin (1988).
Таким образом, на дне, как и на суше, наблюдаемая мозаика сообществ есть отражение сукцессионной динамики, проходящей в гетерогенной среде при постоянных нарушениях. Как и в наземных ЦС, конечным результатом сукцессии в бентосе является формирование климаксного сообщества.
Хотя ассоциации обычно маркируют по крупным долгоживущим видам, которые часто являются эдификаторами, наличие таких видов вовсе не обязательно и сукцессия не сводится к смене таких видов. Другие компоненты сообщества, обладающие более короткой продолжительностью жизни, также сменяются. Смена одного вида другим может происходить только после смерти особей. Поэтому в реальности смена ассоциаций в процессе сукцессии происходит не дискретно-прерывисто, а плавно. Это — одна из причин делающих столь устойчивыми представления о существовании континуума.
Климакс — это ассоциация, наиболее приспособленная к существующим в конкретном месте условиях. На суше климакс обусловлен климатически (климатический климакс). Clements (1936) подчеркивал, что climax и climate имеют один греческий корень. Основной фактор, определяющий распределение наземных ЦС (климаксов) — температура (Разумовский, 1999). Основной фактор, по градиенту которого формируются ассоциации в пределах одной ЦС — влажность, причём влажность в значительной степени контролирует сама ЦС и тем в большей степени, чем ближе ассоциация в сукцессионном ряду к климаксу. Оба фактора варьируют на большей части поверхности суши сравнительно плавно, исключение составляют лишь крупные горные системы.
Самые ранние стадии формирования ЦС мы можем наблюдать на примере городских экосистем. Из-за слабой предсказуемости условий в городах до видообразования дело пока не доходит, дело ограничивается подбором видов, соответствующих доступным ресурсам. Например, в российских городах огромные площади заброшены и неухожены и заросли ценофобами. Одним из основных потребителей их семян являются воробьи. В Лондоне таких площадей почти нет (английские газоны!), соответственно воробьи полностью отсутствуют, как это ни удивительно. Одни и те же ресурсы в разных местах могут потреблять разные виды. В Лондоне, как и крупных российских городах, есть ресурсы для крупного хищного млекопитающего, способного питаться падалью (отбросами). В Москве эти ресурсы потребляют бродячие собаки, берущие начало от брошенных собак. В Лондоне за судьбой домашних питомцев следит специальная служба. Естественно, нет и бродячих собак. Но ресурсы есть, и их потребляют многочисленные дикие лисы. Освоение новой для вида среды обитания происходит не по всему ареалу, а локально. Популяции, освоившие городскую среду, в дальнейшем расселяются по городам, часто не смешиваясь с обитающими в окрестной дикой природе. Так, обитатель глухих лесов вяхирь в Западной Европе перешёл к жизни в городах, вероятно, из-за полного уничтожения глухих лесов. Вяхири с изменённым поведением постепенно расселяются на восток, при этом вяхири в лесах остаются столь же пугливыми, как и ранее: это свидетельствует об отсутствии обменов между городской и природной популяциями. Эти две репродуктивно изолированные популяции, по-видимому, можно рассматривать как разные виды (о видах, причинах их образования и критериях см. ниже в этой Главе).