Учебник Биология - Для учащихся медицинских училищ и колледжей - 2016 год
Теории происхождения протобиополимеров - ВОЗНИКНОВЕНИЕ ЖИЗНИ НА ЗЕМЛЕ
Различные оценки характера среды на первобытной Земле привели к созданию разных условий экспериментов, имевших принципиально единые, но не всегда одинаковые в частностях результаты.
Рассмотрим некоторые из важнейших теорий возникновения полимерных структур на нашей планете, лежащих у истоков образования биополимеров — основы жизни.
Термическая теория. Реакции конденсации, которые привели бы к образованию полимеров из низкомолекулярных предшественников, могут осуществляться путем нагревания. По сравнению с другими компонентами живой материи наиболее хорошо изучен синтез полипептидов.
Инициатором синтеза полипептидов термическим путем является американский ученый С. Фокс, который длительное время изучал возможности образования пептидов в условиях, существовавших на первобытной Земле. Если смесь аминокислот нагреть до 180 — 200 °С при нормальных атмосферных условиях или в инертной среде, то образуются продукты расщепления, небольшие олигомеры, в которых мономеры соединены пептидными связями, а также малые количества полипептидов. Когда исходные смеси аминокислот обогащались экспериментаторами аминокислотами кислого или основного типа, например, аспарагиновой и глутаминовой кислотами, доля полипептидов значительно возрастала. Молекулярная масса полимеров, полученных таким путем, может достигать нескольких тыс. D (D — Дальтон, единица измерения массы, численно равная массе одного атома углерода: 1,67 х 10-24 г. — Прим. ред.).
Полимеры, полученные термическим путем из аминокислот, — протеиноиды — проявляют многие специфические свойства биополимеров протеинового типа. Однако в случае конденсации термическим путем нуклеотидов и моносахаридов, имеющих сложную структуру, образование известных в настоящее время нуклеиновых кислот и полисахаридов представляется маловероятным.
Теория адсорбции. Основным контраргументом в спорах об абиогенном возникновении полимерных структур является концентрационный барьер и недостаток энергии для конденсации мономеров в разбавленных растворах. И действительно, по некоторым оценкам, концентрация органических молекул в “первичном бульоне” составляла около 1%. Такая концентрация в силу редкости и случайности контактов различных молекул, необходимых для конденсации веществ, не могла обеспечить столь быстрого образования протобиополимеров, как это имело место на Земле, по оценкам некоторых ученых. Одно из решений этого вопроса, связанное с преодолением концентрационного барьера, было предложено английским физиком Д. Берналом, считавшим, что концентрирование разбавленных растворов происходит путем “адсорбции в пресноводных или морских отложениях очень тонких глин”.
В результате взаимодействия веществ в процессе адсорбции некоторые связи ослабляются, что приводит к разрушению одних и образованию других химических соединений.
Низкотемпературная теория. Авторы данной теории, румынские ученые К. Симонеску и Ф. Денеш, исходили из несколько иных представлений об условиях абиогенного возникновения простейших органических соединений и их конденсации в полимерные структуры. Ведущее значение в качестве источника энергии авторы придают энергии холодной плазмы. Такое мнение небезосновательно.
Холодная плазма широко распространена в природе. Ученые полагают, что 99% Вселенной находится в состоянии плазмы. Встречается
это состояние материи и на современной Земле в виде шаровых молний, полярных сияний, а также наиболее важного типа плазмы — ионосферы.
Вне зависимости от характера энергии на абиотической Земле любой ее вид преобразует химические соединения, в особенности органические молекулы, в активные частицы, такие, как моно- и полифункциональные свободные радикалы. Однако дальнейшая их эволюция в значительной степени зависит от плотности энергетического потока, который в случае использования холодной плазмы наиболее ярко выражен.
В результате проведения кропотливых и сложных экспериментов с холодной плазмой в качестве источника энергии для абиогенного синтеза протобиополимеров исследователям удалось получить как отдельные мономеры, так и полимерные структуры пептидного типа, и липиды.
Коацерватная теория. Автором этой теории является известный отечественный биохимик академик А.И. Опарин (1924). Несколько позже, независимо от него, к аналогичным выводам пришел английский ученый Дж. Холдейн (1929).
Опарин считал, что переход от химической эволюции к биологической требовал обязательного возникновения индивидуальных фазово-обособленных систем, способных взаимодействовать с окружающей внешней средой, используя ее вещества и энергию, и на этой основе способных расти, множиться и подвергаться естественному отбору.
Абиотическое выделение многомолекулярных систем из однородного раствора органических веществ, по-видимому, должно было осуществляться многократно. Оно и сейчас очень широко распространено в природе. Но в условиях современной биосферы можно непосредственно наблюдать только начальные стадии образования таких систем. Их эволюция обычно очень кратковременна в присутствии уничтожающих все микробов. Поэтому для понимания этой стадии возникновения жизни необходимо искусственно получать фазово-обособленные органические системы в строго контролируемых лабораторных условиях и на сформированных таким образом моделях устанавливать, как пути их возможной эволюции в прошлом, так и закономерности этого процесса. При работе с высокомолекулярными органическими соединениями в лабораторных условиях ученые постоянно встречаются с образованием такого рода фазово-обособленных систем. Поэтому можно представить себе пути их возникновения и экспериментально получить в лабораторных условиях разнообразные системы, многие из которых могли бы послужить моделями возникавших когда-то на земной поверхности образований. Для примера можно назвать некоторые из них: “пузырьки” Гольдейкра, “микросферы” Фокса, “джейвану” Бахадура, “пробионты” Эгами и многие другие.
Часто при работе с такими искусственными самоизолирующимися из раствора системами особое внимание уделяется их внешнему морфологическому сходству с живыми объектами. Но не в этом лежит решение вопроса, а в том, чтобы система могла взаимодействовать с внешней средой, используя ее вещества и энергию по типу открытых систем, и на этой основе расти и множиться, что характерно для всех живых существ.
Наиболее перспективными в этом отношении моделями могут служить коацерватные капли.
Каждая молекула имеет определенную структурную организацию, т.е. атомы, входящие в ее состав, закономерно расположены в пространстве. Вследствие этого в молекуле образуются полюсы с различными зарядами. Например, молекула воды Н2O образует диполь, в котором одна часть молекулы несет положительный заряд (+), а другая — отрицательный (—). Кроме этого, некоторые молекулы (например, соли) в водной среде диссоциируют на ионы. В силу таких особенностей химической организации молекул вокруг них в воде образуются водные “рубашки” из ориентированных определенным образом молекул воды. На примере молекулы NаСl можно заметить, что диполи воды, окружающие ион Na+, обращены к нему отрицательными полюсами, а к иону Сl_ — положительными (рис. 21.3).
Органические молекулы имеют большую молекулярную массу и сложную пространственную конфигурацию, поэтому они тоже окружены водной оболочкой, толщина которой зависит от величины заряда молекулы, концентрации солей в растворе, температуры и др.
При определенных условиях водная оболочка приобретает четкие границы и отделяет молекулу от окружающего раствора. Молекулы, окруженные водной оболочкой, могут объединяться, образуя многомолекулярные комплексы — коацерваты.
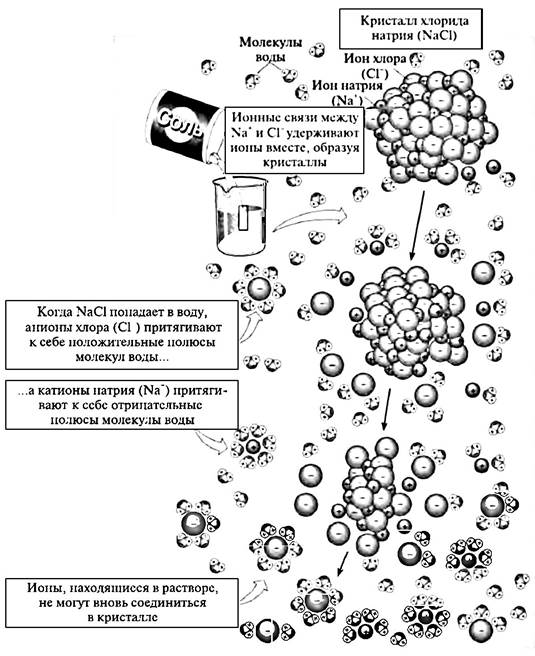
Рис. 21.3. Растворение в воде хлорида натрия и образование гидратированных ионов
Коацерватные капли возникают также при простом смешивании разнообразных полимеров, как естественных, так и искусственно полученных. При этом происходит самосборка (объединение) полимерных молекул в многомолекулярные фазово-обособленные образования — видимые под микроскопом капли (рис. 21.4). В них сосредо-
точивается большинство полимерных молекул, тогда как окружающая среда оказывается почти полностью их лишена.
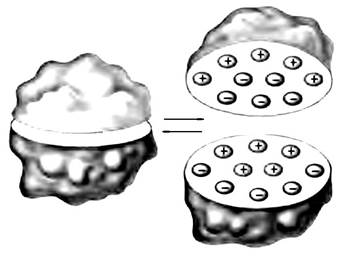
Рис. 21.4. Схема строения коацерватной капли
Капли отделены от окружающей среды резкой границей раздела, но они способны поглощать извне вещества по типу открытых систем.
Путем включения в коацерватные капли различных катализаторов (в том числе и ферментов) можно вызывать ряд реакций, в частности полимеризацию поступающих из внешней среды мономеров. За счет этого капли могут увеличиваться в объеме и весе, а затем дробиться на дочерние образования.
Например, процессы, протекающие в коацерватной капле, изображены в квадратных скобках, а вне их помещены вещества, находящиеся во внешней среде:
глюкозо-1 -фосфат →
[глюкозо-1 -фосфат → крахмал → мальтоза] →
мальтоза.
Коацерватную каплю, образованную из белка и гуммиарабика, погружают в раствор глюкозо-1-фосфата. Глюкозо-1-фосфат начинает входить в каплю и полимеризуется в ней в крахмал при действии катализатора — фосфорилазы. За счет образовавшегося крахмала капля растет, что легко может быть установлено как химическим анализом, так и непосредственными микроскопическими измерениями. Если в каплю включить другой катализатор — b-амилазу, крахмал распадается до мальтозы, которая выделяется во внешнюю среду.
Таким образом, возникает простейшая форма метаболизма. Вещество входит в каплю, полимеризуется, обусловливая рост системы, а при распаде полимера продукты этого процесса выходят во внешнюю среду, где их ранее не было.
Другая схема иллюстрирует опыт, где полимером является полинуклеотид. Капля, состоящая из белка-гистона и гуммиарабика, окружена раствором АДФ.
Поступая в каплю, АДФ полимеризуется под влиянием полимеразы в полиадениловую кислоту, за счет которой капля растет, а неорганический фосфат поступает во внешнюю среду:
АДФ → [АДФ→ Поли-А + Ф] → Ф.
При этом капля в течение короткого срока увеличивается в объеме более чем в два раза.
Как в случае синтеза крахмала, так и при образовании полиадени- ловой кислоты в качестве исходных веществ в окружающий раствор вносили богатые энергией (макроэргические) соединения. За счет энергии этих соединений, поступающих из внешней среды, и происходили синтез полимеров и рост коацерватных капель. В другой серии опытов академика А.И. Опарина и сотрудников было продемонстрировано, что и в самих коацерватных каплях могут протекать реакции, связанные с рассеиванием энергии.
Важно то, что в зависимости от совершенства внутренней организации капель одни из них могут расти быстро, тогда как другие, находясь в той же внешней среде, замедлены в своем росте или подвергаются распаду. Таким образом, на модели коацерватных капель А.И. Опарину и его сотрудникам удалось экспериментально наблюдать зачатки естественного отбора — той закономерности, которая в дальнейшем легла в основу всей последующей эволюции такого рода открытых, фазово-обособленных систем на пути к возникновению жизни.
Белково-коацерватная теория Опарина — одна из первых научных теорий происхождения жизни абиогенным путем. Теория базируется на представлении о том, что все начиналось с белков, а также на возможности в определенных условиях спонтанного химического синтеза мономеров белков (аминокислот) и белковоподобных полимеров (полипептидов) абиогенным путем. Публикация теории стимулировала многочисленные эксперименты в ряде лабораторий мира, показавшие реальность такого синтеза в искусственных условиях. Теория быстро стала общепринятой и необыкновенно популярной.
Все было хорошо продумано и научно обосновано в этой теории, кроме одной проблемы. Если спонтанно, путем случайных безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул (например, эффективные катализаторы, обеспечивающие преимущество данному коацервату в росте и размножении), то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения внутри коацервата и в поколениях единичных, случайно появившихся эффективных белковых структур.
Мир РНК как предшественник современной жизни. Накопление знаний о нуклеиновых кислотах, генетическом коде и биосинтезе белков привело к утверждению принципиально новой идеи о том, что жизнь начиналась вовсе не с белков, а с РНК. Нуклеиновые кислоты являются единственным типом биологических полимеров, макромолекулярная структура которых благодаря принципу комплементарности при синтезе новых цепей обеспечивает возможность копирования собственной линейной последовательности мономерных звеньев, другими словами, возможность воспроизведения (репликации) полимера, его микроструктуры. Поэтому только нуклеиновые кислоты, но не белки могут быть генетическим материалом, т.е. воспроизводимыми молекулами, повторяющими свою специфическую микроструктуру в поколениях.
По-видимому, именно РНК, а не ДНК могла представлять собой первичный генетический материал.
Во-первых, в химическом синтезе и в биохимических реакциях ри- бонуклеотиды предшествуют дезоксирибонуклеотидам; содержащие дезоксирибозу нуклеотиды являются продуктами модификации рибо- нуклеотидов.
Во-вторых, в самых древних и универсальных процессах метаболизма, протекающих в клетках современных организмов, широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители типа АТФ и других.
В-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, механизм же редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза полинуклеотидной цепи ДНК.
В-четвертых, РНК, обладая теми же матричными и генетическими функциями, что и ДНК, способна кроме этого к выполнению ряда функций, присущих белкам, включая катализ химических реакций.
Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение — модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
После того как были открыты каталитически активные РНК, идея первичности РНК в происхождении жизни получила сильнейший стимул к развитию. Американским ученым В. Гилбертом в 1986 г. была сформулирована концепция самодостаточного мира РНК, предшествовавшего современной жизни. Возможная схема возникновения мира РНК представлена на рис. 21.5.
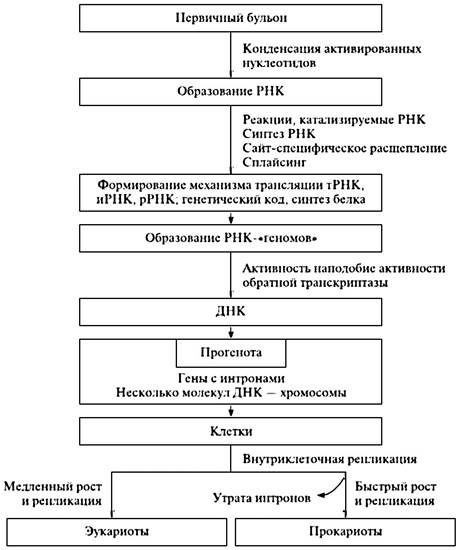
Рис. 21.5. Возможный механизм доклеточной эволюции
Абиогенный синтез рибонуклеотидов и их ковалентное объединение в полимеры РНК могли происходить приблизительно в тех же условиях и в той же химической обстановке, что постулировались для образования аминокислот и полипептидов. Позднее было экспериментально показано, что по крайней мере некоторые молекулы РНК в обычной водной среде способны к спонтанной рекомбинации, т.е. обмену отрезками полинуклеотидной цепи. Обмен коротких отрезков цепи на длинные должен приводить к удлинению полирибо- нуклеотидов (РНК), а сама подобная рекомбинация способствовать структурному многообразию этих молекул. Среди них могли возникать и каталитически активные молекулы РНК — рибозимы.
Даже крайне редкое появление единичных молекул РНК, которые были способны катализировать полимеризацию рибонуклеотидов или соединение олигонуклеотидов на комплементарной цепи как на матрице, означало становление механизма репликации РНК. Создавая свои копии, РНК размножались. Неизбежные ошибки в копировании — мутации и рекомбинации в самореплицирующихся популяциях РНК — создавали все большее разнообразие этого мира. Таким образом, предполагаемый древний мир РНК представляет собой самодостаточный биологический мир, в котором молекулы РНК функционировали и как генетический материал, и как энзимоподобные катализаторы.
Возникновение биосинтеза белка. Впоследствии на основе мира РНК должны были происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, возможно, в форме коацерватов и эволюция этих образований в клеточные структуры.
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру — наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза полипептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белке в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Одна из таких гипотез принадлежит академику А.С. Спирину; она включает два принципиальных момента.
Во-первых, предполагается, что олигорибонуклеотиды, образующиеся абиогенным путем, активно рекомбинировали, приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем среди полинуклеотидов и могли появиться как каталитически активные виды РНК — рибозимы, так и другие виды РНК со специализированными функциями (см. рис. 21.5). Более того, неферментная рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов могло осуществляться первичное копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью, то эффективность (точность, скорость и продуктивность) копирования на комплементарной матрице значительно возрастала.
Второй принципиальный момент состоит в том, что первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК. Этот первичный аппарат включал каталитически активную прорибосомную РНК; набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды; другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-иРНК и про-тРНК (см. рис. 21.5). Такая система уже могла синтезировать полипептидные цепи. Среди прочих каталитически активных белков — первичных ферментов (энзимов) появились и белки, катализирующие полимеризацию нуклеотидов, — репликазы, или полимеразы.
Таким образом, РНК представляется наиболее самодостаточным веществом живой материи. Она принципиально способна выполнять все или почти все функции, которые свойственны белкам, включая формообразование и биохимический катализ, и в то же время может быть полноценным генетическим веществом с его репликативной и кодирующей функциями. Осознание этих фактов и привело биологов, химиков и геологов к гипотезе о древнем “мире РНК”, который эволюционно предшествовал нашей нынешней ДНК-РНК-белковой жизни. В мире РНК не было ни белков, ни ДНК, а лишь ансамбли различных молекул РНК, выполняющих разные вышеперечисленные функции. Это были, скорее всего, бесклеточные системы. Около десяти лет назад у нас в стране была экспериментально показана способность молекул РНК формировать молекулярные колонии на гелях или других твердых средах. Смешанные колонии РНК на твердых или полутвердых поверхностях могли быть первыми эволюционирующими бесклеточными ансамблями, где одни молекулы выполняли генетические функции, а другие формировали необходимые для успешного существования структуры (например, такие, которые адсорбировали нужные вещества из окружающей среды) или были рибозимами, ответственными за синтез и подготовку синтеза РНК. Такая бесклеточная ситуация создавала условия для очень быстрой эволюции: колонии РНК не были отгорожены от внешней среды и могли легко обмениваться своими молекулами. Легкое распространение молекул РНК через среду, в том числе атмосферную, также было продемонстрировано в прямых экспериментах.
Именно такие условия были необходимыми для возникновения Универсального Предшественника живых существ на Земле: высокий уровень мутаций (ошибок репликации) из-за примитивности и несовершенства механизмов репликации генетического материала, свободный обмен генетическим материалом между предшественниками клеток — протоклетками — и коммунальный характер существования этих предшественников, когда любые продукты одних становились достоянием всех.
По мнению академика А.С. Спирина, эта коммунальная форма существования мира РНК — своего рода Солярис — должна была очень быстро эволюционировать. Во всяком случае весь путь эволюции до индивидуальных организмов с клеточной структурой, ДНК и современным аппаратом белкового синтеза был пройден, по-видимому, менее чем за 500 млн лет. Совершенствование колоний-ансамблей РНК за счет естественного отбора должно было происходить в направлении как улучшения каталитических механизмов, так и увеличения точности репликации и наследования. Колонии РНК, “научившиеся” делать белковые катализаторы, естественно, приобретали громадное преимущество перед другими в скоростях и качестве катализируемых реакций и потому быстро вытесняли “неумелых” — как за счет конкуренции, так и за счет передачи им этой способности. На базе РНК появлялся и совершенствовался аппарат белкового синтеза, а ввиду коммунального и пандемического характера мира РНК вырабатывался универсальный генетический код.
Однако кодируемый синтез белков требовал повышенной точности репликации генетического материала и упорядочивания продукции разных белков. Это привело к необходимости дифференциации части РНК (генетической РНК) и ее модификации в ДНК, обладающую способностью к более точному копированию, а к тому же и существенно большей химической стабильностью, чем РНК. Наконец, эффективность и устойчивость таких систем могла быть значительно повышена за счет их обособления от окружающей среды, и они окружаются мембранами белково-липидной природы. Коммунальный мир распадается на индивидуальные, но высокоэффективные ячейки — клетки, особи, организмы, и начинаются их собственная эволюция и собственные родословные. Из коммунального Универсального Предшественника выходят две основные ветви микроорганизмов — бактерии (эубактерии) и археи (архебактерии), формируются их клеточные сообщества на основе взаимодействия их метаболизма, а затем их симбиотические отношения приводят к появлению эукариотических организмов.
Опорные точки
• Органические вещества по отношению к воде подразделяются на две крупные группы: гидрофобные и гидрофильные молекулы.
• В водных растворах гидрофильные молекулы диссоциируют, образуя заряженные частицы.
• Крупные органические молекулы, обладающие зарядом, либо связываются с субстратом, либо взаимодействуют друг с другом, в результате чего формируются коацерваты.
• Рибонуклеиновые кислоты обладают способностью к специфическому катализу, самовоспроизведению и другим формам матричного синтеза.
Вопросы и задания для повторения
1. Что такое коацервация, коацерват?
2. На каких модельных системах можно продемонстрировать образование коацерватных капель в растворе?
3. Какие возможности для преодоления низких концентраций органических веществ существовали в водах первичного океана?
4. Что собой представляет “мир РНК” и каково его значение для начальных этапов эволюции жизни на Земле?
5. В чем заключаются преимущества для взаимодействия органических молекул в зонах высоких концентраций веществ?
6. Как вы думаете, могут ли в современных земных условиях образовываться небиологическим путем органической молекулы? Коацерваты?
7. Как, по вашему мнению, происходило становление системы синтеза белков у протобионтов?
8. Каковы перспективы эволюции коацерватов, полученных экспериментальным путем, в окружающей среде?