БИОЛОГИЯ пособие для поступающих в вузы
Том ІІ ботаника, анатомия и физиология, эволюция и экология 2018 год
Учение Дарвина об отборе легло в основу такой науки, как селекция. Предметом селекции является исследование специфических закономерностей эволюции домашних животных и возделываемых растений, направляемой человеком. Главная цель и задачи селекции состоят в создании высокопродуктивных пород животных и сортов растений, которые наилучшим образом повышают производительность труда в животноводстве и растениеводстве. Порода и сорт входят в средства сельскохозяйственного производства. Породой и сортом называют популяцию организмов, искусственно создаваемую человеком и имеющую определенные наследственные особенности. Все особи внутри породы и сорта имеют очень сходные, наследственно закрепленные показатели продуктивности, биологические свойства и морфологические признаки.
Основные методы селекции
1. Генетические методы. Изменчивость исходного материала является основой для создания новых пород животных и сортов растений. При этом имеют значение такие типы изменчивости, как а) мутационная', б) комбинационная; в) полиплоидия.
2. Методы скрещивания. Наличие наследственной изменчивости позволяет путем различных систем скрещивания сочетать определенные наследственные признаки в одном организме, а также избавляться от нежелательных свойств. При разведении применяют различные системы скрещиваний, которые делят на несколько типов:
а) родственное скрещивание, часто называемое инбридин гом. Родственным называют скрещивание особей, имеющих близкую степень родства: брат—сестра, двоюродные братья и сестры и т. д. Его смысл заключается в том, что генетический состав популяции оценивается путем разложения ее на генетически различные линии. При этом гены, находящиеся в гетерозиготном состоянии, переходят в гомозиготное состояние;
б) неродственное скрещивание, когда у скрещиваемых особей имеется отсутствие ближайших общих предков в 4 — 6 поколениях их родословных. Ценность этого скрещивания в том, что помеси первого поколения оказываются более жизнеспособными, более устойчивыми к заболеваниям, имеют повышенную плодовитость, т. е. проявляют гетерозис, или явление гибридной мощности. Под гетерозисом понимают свойство гибридов первого поколения при скрещивании различных рас, пород животных и сортов растений, а также инбридных (чистых) линий превосходить по ряду признаков и свойств лучший из исходных родительских организмов. Дальнейшее скрещивание гибридов между собой ведет к затуханию этого эффекта в следующих поколениях. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только родителей (линии), но и сорта или породы, от которых произошли эти линии. В настоящее время гетерозис установлен для всех изученных видов.
3. Методы отбора. Отбор является одним из основных методов селекции. В сочетании с генетическими методами он позволяет создавать новые формы, сорта и плоды с определенными свойствами и признаками. Различают:
а) массовый отбор — т. е. отбор особей по внешним показателям (фенотипу), отвечающим принятому стандарту для популяции сорта или породы без проверки генотипа отдельных особей;
б) индивидуальный отбор — т. е. отбор, при котором прежде всего оценивается потомство отдельного растения или животного в ряду поколений. Вследствие этого становится возможным оценивать наследственные качества отдельных индивидуумов — способность передавать свои свойства и норму реакции генотипа потомству.
4. Онтогенетический метод. Крупный вклад в развитие селекции растений внес русский ученый И. В. Мичурин. Им поднят совершенно новый для селекционно- генетической теории вопрос, а именно проблема развития наследственных признаков в онтогенезе. Он доказал, что сортовые признаки растений, например, их теплолюбивость или холодоустойчивость, форма, размер и качество плодов, формируются на ранних стадиях онтогенеза под влиянием наследственности и условий внешней среды. И. В. Мичурин создал стройную систему селекционного процесса, в которой сочетаются три основных звена: гибридизация, воздействие условиями внешней среды на развивающиеся гибриды и отбор. Мичурин разработал специальные методы преодоления нескрещиваемости у растений при отдаленной гибридизации. (Отдаленной гибридизацией называется скрещивание форм, относящихся к разным разновидностям, видам, родам и другим таксономическим единицам более высокого порядка. Например, мул был получен при скрещивании лошади и осла; грейпфрут — лимона и апельсина.)
К числу основных методов Мичурина относятся:
а) метод предварительного вегетативного сближения. Заключается в предварительной прививке скрещиваемых растений друг на друга (например, сеянец рябины гибридного происхождения прививают в крону взрослого дерева груши для дальнейшего преодоления нескрещиваемости между рябиной и грушей);
б) метод посредника. Данный метод лучше всего рассмотреть на конкретном примере. Мичурин, чтобы продвинуть на Север культуру персика, решил осуществить гибридизацию этого культурного растения с диким монгольским миндалем бобовником. Поскольку прямого скрещивания между ними добиться не удалось, экспериментатор предварительно скрестил миндаль с полукультурным персиком Давида. Полученный от скрещивания гибрид Мичурин назвал посредником, так как он уже сравнительно легко скрещивался с культурными сортами персика;
в) метод направленного воспитания сеянцев. Здесь особенно известен метод “ментора”. “Ментором” Мичурин называет взрослое растение, которое служит воспитателем для молодого сеянца;
г) метод опыления смесью пыльцы в целях преодоления нескрещиваемости растений. Так, при скрещивании яблони (мать) с грушей (отец) к пыльце последней добавляется небольшое количество пыльцы яблони. Это делает рыльце яблони восприимчивым и к пыльце груши, вследствие чего часть семяпочек оплодотворяется материнской пыльцой, а часть — пыльцой чужого вида.
Селекция и учение Н. И. Вавилова
Процесс окультуривания растений имеет разную длительность: у одних он продолжается несколько тысячелетий, а у других начался не более двухсот лет назад. В решении проблемы происхождения культурных растений ведущую роль сыграло учение академика Н. И. Вавилова о центрах происхождения культурных растений. В результате его исследований было установлено семь очагов земного шара, откуда ведет начало вся мировая земледельческая культура (табл. 6.2, рис. 6.45).
Рис. 6.45. Центры происхождения культурных растений
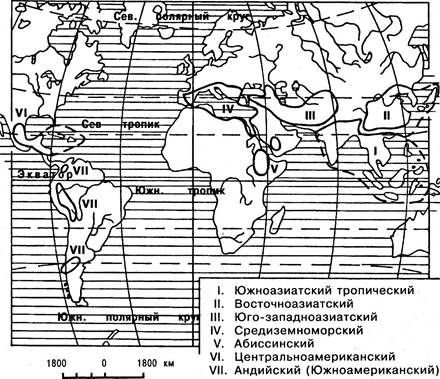
Таблица 6.2
Центры происхождения культурных растений
|
Центры |
Растения |
|
1. Южноазиатский |
Тропический рис, сахарный тростник, значительная часть зерновых бобовых, тропические плодовые — банан, кокосовая пальма |
|
2. Восточноазиатский |
Просо, гречиха, ряд клубнеплодов и корнеплодов, многие плодовые (груша, яблоня, слива и др.), некоторые цитрусовые и декоративные растения |
|
3. Юго-западноазиатский |
Мягкая пшеница, карликовая пшеница, круглозерная пшеница, горох, чечевица, чина, конские бобы, нут, хлопчатник-гуза |
|
4. Средиземноморский |
Маслина, рожковое дерево, ряд овощных (свекла и др.) |
|
5. Абиссинский |
Твердая пшеница, ячмень, кофейное дерево |
|
6. Центральноамериканский |
Кукуруза, американские фасоли, тыквы, перец, какао, хлопчатник- упланд |
|
7. Андийский (Южноамериканский) |
Картофель и ряд других корнеплодов, табак, ананас, земляной орех, хинное дерево |
Детальное изучение изменчивости многих видов растений дало возможность Н. И. Вавилову обобщить все известные факты в форме общего закона, известного под названием закона гомологичных рядов в наследственной изменчивости. Сущность этого закона заключается в следующем: “генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов”.
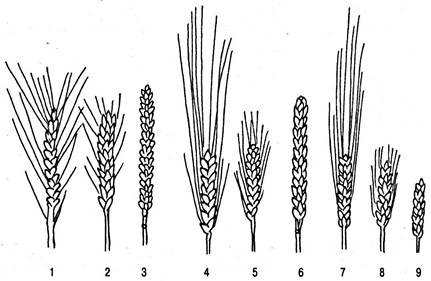
Закон дает возможность обнаружить сходную направленность изменчивости, например, у злаковых (рис. 6.46), и предугадать существование форм, еще не известных науке.
Рис. 6.46. Гомологические ряды изменчивости у видов пшеницы и ячменя по признаку остистости: 1 — 3 — формы мягких пшениц (42 хромосомы); 4 — 6 — формы твердых пшениц (28 хромосом); 7 — 9 — формы шестирунного ячменя (1,4, 7 — остистые формы; 2, 5, 8 — короткоостые формы; 3, 6, 9 — безостые формы)
Факторы эволюции
I. Изоляция как элементарный фактор эволюции. Изоляция — возникновение любых барьеров, препятствующих скрещиванию данного вида с другими видами. Различают два вида изоляции.
1. Пространственная, или географическая, изоляция. Эта изоляция может существовать в разных формах: водные барьеры разделяют “сухопутные” виды, барьеры суши изолируют виды-гидробиоты, возвышенности — равнинные популяции, а равнины — горные популяции и т. д.
2. Репродуктивная изоляция. Предотвращает межвидовые скрещивания, а также уменьшает успешность межвидовых скрещиваний. Это связано с отсутствием встреч между потенциально брачными партнерами (сезонная, или биотопическая, изоляция); с отсутствием спаривания при встрече между потенциально брачными партнерами (этологическая изоляция); с отсутствием осеменения при попытке к копуляции (механическая изоляция); с нежизнеспособностью оплодотворенных яиц либо неполноценностью последующих поколений (гибридологическая изоляция).
II. Изменчивость как элементарный фактор эволюции. Без изменения генотипического состава популяции невозможно протекание любого эволюционного процесса. Генетическая гетерогенность популяции связана с качественными перестройками генофонда. С генетической точки зрения, эволюция —
это изменение генов в данной популяции. Частота какого- либо гена (точнее, аллели) зависит от ряда различных факторов, среди которых следует отметить:
1. Мутационный процесс. Мутация — это изменение, затрагивающее последовательность нуклеотидов или их число в молекуле ДНК и создающее тем самым новые аллели.
2. Рекомбинация. Процесс, с помощью которого имеющаяся в популяции аллель перемешивается (рекомбинирует) в результате скрещивания между собой. Эта генетическая комбинаторика многократно изменяет значение мутаций: они входят в новые геномы, оказываются в новой генотипической среде.
3. Поток генов. Частота генов в данной популяции может изменяться в результате эмиграции или иммиграции особей. Это обмен генами между популяциями, осуществляющийся в форме происходящих время от времени спариваний между особями, принадлежащими к различным скрещивающимся популяциям, проникновения в данную популяцию животного, родившегося в другом месте, или же внесения чужих семян или пыльцы. Непосредственный эффект такого события сходен с эффектом мутации — в популяцию вносится новый источник изменчивости.
4. Закон Харди — Вайнберга и поддержание изменчивости.Английский математик Г. Харди и независимо от него немецкий генетик В. Вайнберг сформулировали положение, которое сейчас именуется законом Харди — Вайнберга и описывает частоту встречаемости генов в равновесной популяции свободно скрещивающихся организмов. Этот закон описывает некую абстрактную “идеальную” популяцию со свойствами, для реальных популяций невозможными:
а) численность такой популяции должна равняться бесконечности;
б) скрещивание особей внутри популяции полностью свободно, т. е. осуществляется без всяких ограничений. Такие популяции называются панмиксными (свободно смешивающимися);
в) популяция должна быть абсолютно изолированной.
Допустим, что частота встречаемости доминантного гена А в популяции равна g, а рецессивного а соответственно 1 - g, тогда соотношение гомо- и гетерозигот будет следующим:
g2АА + 2g (1 — g)Аа + (1 — g)2аа
Это известная из школьной алгебры формула квадрата суммы двух чисел. Если g = 0,5, как в случае первого поколения менделевских гибридов, то во втором поколении мы имеем следующее соотношение:
0,25 АА + 0,5 Аа + 0,25 аа,
а так как Аа в случае полной доминантности фенотипически выражены как АА, мы получаем менделевское расщепление — 0,75:0,25, т. е. 3: 1.
5. Дрейф генов (генетико-автоматические процессы). Итак, относительная численность аллелей в популяции абсолютно точно описывалась бы уравнением Харди — Вайнберга, если бы численность самой популяции была бы бесконечной. Чем меньше популяция, тем больше отклонения, и, если численность размножающихся особей в ней окажется ниже 500, начинаются любопытные явления. Их назвали генетико-автоматическими процессами, или коротко, дрейфом генов. Суть дрейфа генов в том, что из-за случайных колебаний один из аллелей может вообще выпасть из популяции, и она окажется по данному гену гомозиготной. В результате происходит гомозиготизация особей и затухание изменчивости. Нетрудно понят]., что это открывает путь случайной дифференцировке популяций вплоть до видового обособления.
III. Популяционные волны как элементарный эволюционный фактор. Популяционными волнами, или “волнами жизни” (по С. С. Четверикову), называют периодические или апериодические колебания численности организмов в природных популяциях. Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. При росте численности организмов наблюдаются слияние ранее разобщенных популяций и объединение их генофондов. При спаде численности наблюдается распад крупных популяций. Возникающие малочисленные популяции характеризуются измененными генофондами.
В условиях массовой гибели организмов редкие мутантные аллели могут быть генофондом вида. При сохранении редкого аллеля его концентрация в генофонде малочисленной популяции автоматически возрастает. Популяционные волны действуют на генофонды ненаправленно, поэтому они так же, как и мутационный процесс, только предоставляют эволюционный материал к действию естественного отбора.
IV. Естественный отбор — движущий фактор эволюции. Под естественным отбором нужно понимать избирательное (дифференциальное) воспроизведение генотипов (или генных комплексов). В процессе естественного отбора важны не столько выживание или гибель особей, сколько их дифференциальное
размножение. Успех в размножении разных особей может служить объективным генетико-эволюционным критерием естественного отбора.
В зависимости от результата различают направленную, стабилизирующую и дизруптивную формы естественного отбора (рис. 6.47).
Рис. 6.47. Воздействие отбора трех различных типов на генетическую изменчивость в популяции
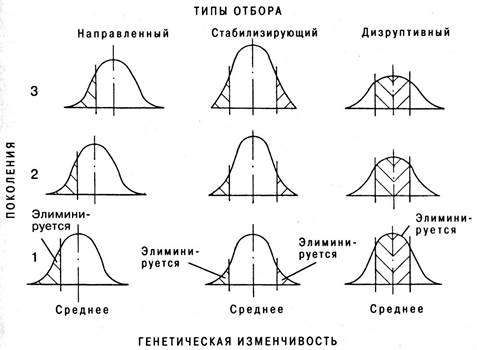
1. Направленный отбор (движущий) вызывает прогрессивное или направленное в одну сторону изменение генетического состава популяций, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. Он происходит в тех случаях, когда популяция находится в процессе приспособления к новой среде или же когда происходит постепенное изменение среды, а вслед за ней и постепенное изменение популяции.
2. Стабилизирующий отбор сохраняет в популяции средний вариант фенотипа или признака. Действие отбора состоит в элиминации плохо приспособленных и периферических вариаций, возникающих в результате мутационного процесса, потока генов и рекомбинации. Это наиболее распространенный тип отбора.
3. Дизруптивный отбор благоприятствует сохранению крайних типов и элиминации промежуточных. В результате он приводит к сохранению и усилению полиморфизма.
Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения в соотношении аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направленны. В эволюции естественному отбору принадлежит творческая роль. Исключая из репродукции генотипы с малой приспособительной ценностью, сохраняя благоприятные генные комбинации разного достоинства, он преобразует картину генотипической изменчивости, складывающуюся первоначально под действием случайных факторов, в биологически целесообразном направлении.
Плата за отбор. Генетический груз. Естественный отбор имеет как позитивный, так и негативный аспекты. Он влечет за собой предпочтительное выживание и размножение одних генотипов и предпочтительную элиминацию, и невозможность размножения других. Невозможность размножения той или иной особи, обусловленная отбором в его негативном аспекте — проявляется ли она в форме избирательной смертности или неудавшегося размножения как такового, известна под названием генетической гибели (или гибели, обусловленной отбором). Генетическая гибель снижает репродуктивный потенциал данной популяции. Популяция обладала бы более высокой численностью, если бы не потери за счет генетической гибели. Поэтому генетическая гибель представляет собой, по крайней мере потенциально, некое бремя для популяции. Это бремя называется генетическим грузом. Различают несколько типов генетического груза, коррелирующегося с разными типами отбора.
1. Мутационный груз — неизбежный побочный продукт мутационного процесса. Этот процесс порождает вредные мутации, которые затем должны быть выметены стабилизирующим отбором.
2. Сегрегационный груз существует в популяциях, которые используют преимущества, создаваемые превосходством гетерозигот. При этом в каждом поколении происходит выщепление менее приспособленных гомозигот, понижающих среднюю приспособленность популяции. Гомозиготы представляют собой плату за уравновешивающий отбор.
3. Субституционный груз, который порождает направленный отбор. Замещение старой аллели, превосходящей ее новой влечет за собой генетическую гибель носителей старой аллели.
Пути и способы видообразования. Процесс образования видов осуществляется в результате взаимодействия элементарных эволюционных факторов. Различают следующие способы видообразования.
1. Аллотропическое видообразование (географическое) — новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии ареала исходного вида.
2. Симпатрическое видообразование — новый вид возникает внутри ареала исходного вида.
3. Дивергентное видообразование — первоначальный единый вид разделяется на два или более новых в связи с возникновением межпопуляционных изоляционных барьеров и углублением различий под действием естественного отбора вплоть до генетической изоляции. Такой процесс, ведущий к увеличению количества видов, называется истинным видообразованием.
4. Филитическое видообразование — новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени без какой-либо дивергенции исходных групп, что наблюдается при изменении условий во всем ареале.
5. Гибридогенное видообразование — новый вид возникает путем гибридизации с последующим удвоением числа хромосом (аллополиплоидия).
Основные направления эволюционного процесса
Изучение филогенеза животного мира показало, что эволюция идет различными путями (рис. 6.48). Вопрос об основных направлениях эволюционного процесса был разработан академиком А. Н. Северцовым.
Рис. 6.48. Схема основных направлений эволюционного процесса. Две верхние плоскости (а, б) возникли путем ароморфоза, нижняя (в) — путем морфологического регресса. Ветвящиеся линии в пределах плоскости — идиоадаптации
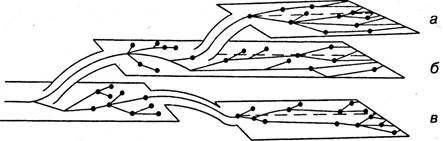
Ароморфоз — основной путь прогрессивной эволюции. В этом направлении шла эволюция от одноклеточных к многоклеточным, от двухслойных к трехслойным, от низших хордовых к позвоночным. Ароморфозы — это подъем организации с одного уровня на другой, более высокий.
Идиоадаптация — путь узкоприспособительных изменений, не связанных с повышением уровня развития. Путь идиоадаптации объясняет возникновение в пределах каждого типа и класса многочисленных видов, имеющих одинаковый уровень организации, но приспособленных, например, к жизни в пресной или в соленой воде.
Морфофизиологический регресс выражается в упрощении строения. Этот путь эволюции, ведущий от сложного к более простому, довольно широко распространен в природе и не приводит к вымиранию упростившихся форм, если они достаточно приспособлены к среде обитания. Примером может служить эволюция ленточных червей, утративших в процессе филогенеза органы пищеварения.