Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Паратетис - Причины и скорость образования новых видов и экосистем - Био-экология, или синэкология
Для Каспийской и Аральской биот и биот распреснённых районов Азово-Черноморского бассейна (и некоторых пресных и солоноватых водоёмов Европы) характерен высокий эндемизм. Данные биоты — остатки биоты Паратетиса.
История Паратетиса вкратце такова. Сближение Индийского субконтинента с Азией к концу олигоцена приводит к полному исчезновению восточной части моря Тетис. Происходившие в то же время горообразовательные процессы отделили от западной части Тетиса Паратетис. В раннем олигоцене обособление Паратетиса привело к его значительному опреснению и вымиранию морской фауны Тетиса. Позднее связь Паратетиса и Тетиса восстанавливается и солоноватоводную биоту Паратетиса сменяет морская биота Тетиса. Сменяющиеся распреснения-осолонения происходили потом многократно. В сарматский век образуется почти замкнутое солоноватоводное Сарматское море. В этом озере-море развивается своеобразная солоноватоводная биота. Сарматское море сменило Мэотическое море с нормальной солёностью. Солоноватоводная биота опять сменилась морской. В конце миоцена Мэотическое море сменяет солоноватоводный Понтический бассейн, который распадается на три основных бассейна: Паннонский, Эвксинский (Чёрноморский) и Каспийский. Причиной этого распадения были как продолжающиеся горообразовательные процессы, так и высыхание Средиземного моря (Мессинский кризис), сброс воды в котловину которого привёл к общему падению уровня воды в Понтическом бассейне. История этих бассейнов была различной. Паннонский ныне почти исчез, его остаток — озеро Балатон, Чёрное море было по преимуществу морским, в Каспии же продолжалась череда опреснений-осолонений. К середине плиоцена (балаханский век) он превращается в гиперсолёное Балаханского озера, заполнявшее лишь впадину южного Каспия. Впадавшие в Балаханское озеро реки (в частности, палео-Волга) прорезали себе в толще пород глубокие и узкие ущелья глубиной несколько сотен метров, а их дельтовые отложения сформировали нефтеносные горизонты тощиной до 4 км. В Акчагыльском веке формируется Акчагыльское море, сообщающееся на севере с Чёрным. После короткой регрессии в начале четвертичного периода возникает Апшеронское море, существенно превосходившее современное Каспийское.
“По мнению К.А. Киселевича (1923), многочисленные виды и подвиды каспийских сельдей произошли от одного вида, а G.O. Sars (1927) и А.Н. Державин (1951) пришли к выводу, что все каспийские Cumacea (только в Каспии 7 родов и 17 видов — ИЖ) произошли от одной прародительской формы” (Карпинский, 2002: 36). По моему мнению все каспийские полихеты- Ampharetidae произошли от одного вида.
Палеонтолог Л.Ш. Давиташвили, характеризуя фауну моллюсков Сарматского века Понто-Каспия, пишет: “Здесь мы видим множество... разнообразнейших форм, многие из них нам пришлось бы выделить в особые роды, если бы мы следовали примеру зоологов, изучающих современную фауну моллюсков” (Давиташвили, 1933: 15(8).
“Эволюционные преобразования фаун замыкавшихся и полностью замкнутых полуморских и солоноватоводных бассейнов могут служить примером преобразования экосистем островного типа, в которых интенсивный формо- и видообразовательный процесс происходил в основном симпатрическим путем и шёл главным образом за счет освоения освобождавшихся экологических ниш. Этот переход в новые, несвойственные для исходной группы ниши, приводил к образованию кроме многочисленных видов таксонов достаточно высокого ранга — родового и семейственного” (Невесская и др., 1987).
Современное население этого района образует три биоты, возникшие из морской биоты Паратетиса и описанные (см. Главу 5): пресноводную (содержит и элементы другого происхождения), солоноватоводную и “морскую”. Пресноводная биота сильно обогатила фауну рек бассейна в первую очередь Волги, а также рек АзовоЧерноморского бассейна: Dreissena, Ampharetidae, Corophiidae и др. Солоноватоводная биота населяет Северный Каспий и существует в виде рефугиумов в устьях рек Азово-Черноморского бассейна, отдельные представители обитают в рефугиумах в других солоноватоводных водоёмах от Марокко до Балтийского моря. “Морская” фауна вместе с несколькими недавними средиземноморскими вселенцами (Cerastoderma, Mytilaster, Abra, Nereis и др.) образовала “морскую” экосистему Среднего и Южного Каспия и Арала.
Л.А. Невесская с соавт. (2006) проанализировали развитие фаун моллюсков в Паратетисе с раннего олигоцена до настоящего времени (за последние 35 млн. лет) (Рис. 1.24). Каждый раз при повышении солёности (что всегда было связано с установлением связи с океаном) биота Паратетиса становилась сходной с биотой районов, с которыми устанавливалась связь. При снижении солёности, вызванной частичной или полной изоляцией, происходила вспышка видообразования. За анализируемый период зарегистрировано 7 таких вспышек, в каждой из которых за короткий период времени возникало большое число видов (десятки и сотни) и родов (десятки). Среди двустворчатых моллюсков это особенно было характерно для Limnocardiinae, куда относится и Cerastoderma — исходный для подсемейства род. Только в данном подсемействе за проанализированный период образовалось более 700 видов 68 родов.
Видообразование во время вспышек происходило гораздо интенсивнее: только в Didacna в плейстоценовом Понто-Каспии за 600 тыс. лет образовалось 70 видов. В других частях Паратетиса вспышки давали другие таксоны: в Паннонском озере (западная часть Паратетиса) в миоцене—плиоцене только Dreissenidae было 130 видов (Geary et. al., 2000). Во всех случаях видовой состав перед вспышками видообразования был очень беден, что с экологических позиций означает наличие неиспользуемых ресурсов.
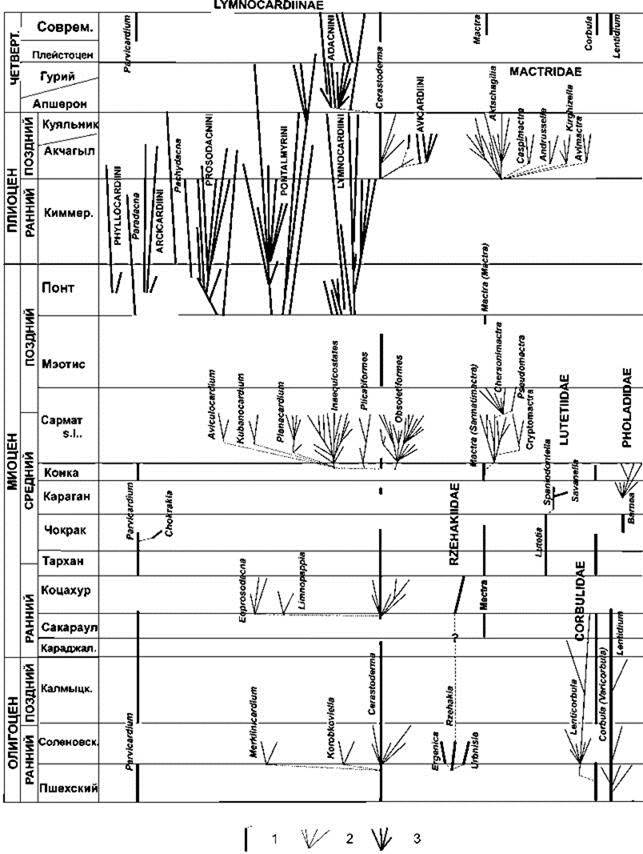
Рис. 1.24. Схема филогении и стратиграфического распространения основных семейств и родов двустворчатых моллюсков, давших вспышки видообразования в бассейнах Восточного Паратетиса По Л.А. Невесской и др. (2006).
1 — исходный морской род; 2 — появление эндемичных видов; 3 — появление эндемичных родов.
В периоды интенсивного видообразования появлялись роды, сильно уклонявшиеся от исходных форм. Cardiidae (Limnocardiinae) становились эпибентосными фильтраторами, одни из них лежали на грунте вниз макушкой, сильно сдвинутой вперёд, другие переходили к жизни на жидких илах, их раковины становились тонкими, на раковинах снаружи могли возникать шипы, помогавшие держаться на поверхности грунта, третьи прикреплялись биссусом к рифам. Дрейссены — ныне исключительно эпибентические фильтраторы — образовывали инфаунных детритофагов (Рис. 1.25). Даже такая узкоспециализированная группа как камнеточцы фоласы в Конкский век дали виды, живущие свободно на мягких грунтах (Давиташвили, 1933). Появление адаптаций, не очень характерных для остальных видов данного таксона, G.S. Myers (1960) назвал “supralimital specialization”, А.И. Кафанов (1991) — “запредельной специализацией”. Очевидно, что в таких случаях происходит попытка освоения нового адаптивного пространства, т. е. ароморфоза.
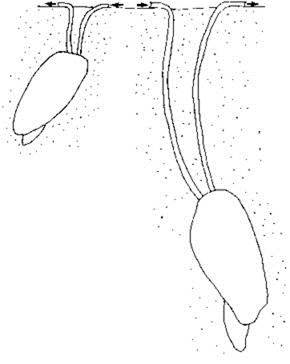
Рис. 1.25. Ископаемые Dreissenidae — Dreissenomya. По Geary et al. (2000).
Представители этого рода известны в отложениях Крыма и Кавказа.
Представление о том, как и с какой скоростью происходило видообразование в водоёмах Паратетиса, даёт изучение современных изменений биоты Аральского моря.