Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Особенности биогеографии континентальных водоёмов - Биогеографическое районирование - Биогеография континентальных водоёмов
Хотя в обзорах, монографиях и учебниках по общей биогеографии биогеографию континентальных водоёмов обычно рассматривают лишь вскользь, опубликовано достаточно много схем их биогеографического районирования (Рис. 4.9—4.12). Правда, все они основаны на анализе таксономического состава. Анализ большинства ЦС континентальных водоёмов — дело будущего. Это связано с объективной сложностью его проведения. Причины этого — во-первых, гораздо большая, чем в морских и наземных ЦС, частота и распространённость экзогенных нарушений. Во- вторых, к континентальным водоёмам относят водоёмы с резко различающимися условиями: крупные реки и озёра, родники, ручейки, речушки, подземные воды, лужи и воды в пазухах листьев или в кувшинах Nepenthes и т. д., и т. п. Многие из них, например, водоёмы в пазухах листьев бромелиевых, мелкие ручьи и лужи эфемерны, а, следовательно, населены по преимуществу или исключительно ценофобами. Поэтому следует ожидать, что доля ценофобных видов в фаунах континентальных водоёмов разных регионов значительно выше, чем в ЦС бентали и суши и даже пелагиали. Большая доля ценофобных видов создаёт шум мешающий познанию биогеографии континентальных водоёмов. Это затрудняет познание закономерностей сукцессий и особенностей функционирования пресноводных водоёмов как единых экосистем различных районов Земли. Однако биоты различных регионов сравнивают тотально, не делая различий по типам водоёмов, биотическим взаимоотношениям и т. д., и т. п. Результаты биотического районирования основаны на распространении всех видов, как ценофильных, так и ценофобных. Районирование, основанное на выделении ЦС проведено только для некоторых крупных озёр (Байкал, Танганьика, Ньяса).
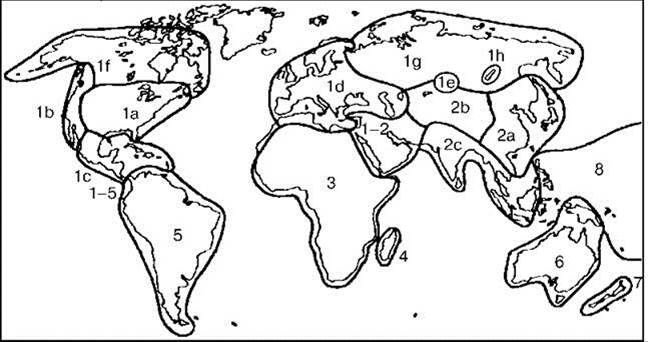
Рис. 4.9. Схемы биогеографического районирования пресных вод P. Banarescu (1992).
Области (подобласти): 1 — Голарктическая (1a — Восточная североамериканская, 1b — Западная североамериканская, 1c — Центральномексиканская, 1d — Евро-средиземноморская, 1e— Западная монгольская, 1f — Арктическая североамериканская, 1g — Сибирская, 1h — Байкальская); 2 — Индо-китайская (2a — Восточноазиатская, 2b — Высокогорная азиатская, 2с — Южноазиатская); 1—2 — Западноазиатская переходная; 3 — Африканская; 4 — Мадагаскарская; 5 — Южноамериканская; 1—5 — Центрально-американскоантильская переходная; 6 — Австралийская; 7 — Новозеландская; 8 — Индо-западнотихоокеанская.
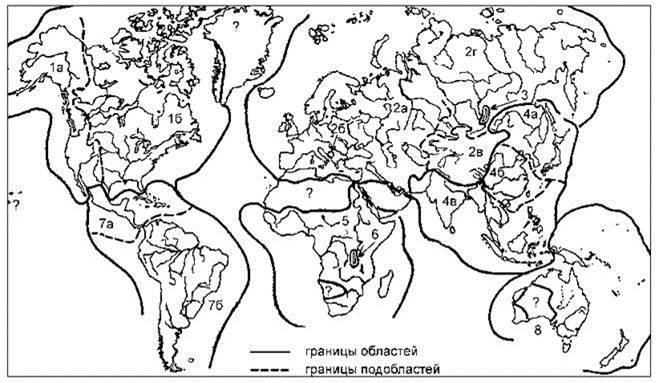
Рис. 4.10. Схема биогеографического районирования пресных вод Я.И. Старобогатова (1970) по распространению моллюсков.
Области (и их подобласти): 1 — Неарктическая (а — Тихоокеанская, б — Атлантическая); 2 — Палеарктическая (а — Европейско-Сибирская, б — Охридская, в — Нагорноазиатская, г — Восточносибирская); 3 — Байкальская; 4 — Сино-Индийская (а — Амуро-Японская, б — Китайская, в — Индо-Малайская); 5 — Эфиопская; 6 — Танганьикская; 7 — Неотропическая (а — Цельтральноамериканская, б — Южноамериканская); 8 — Австралийская. Ранг регионов присвоен на основании значений индекса Престона, т. е. фактически не обоснован.
На карте не показана выделяемая Я.И. Старобогатовым Каспийская солоноватоводная область, поскольку здесь она отнесена к объектам биогеографии моря.
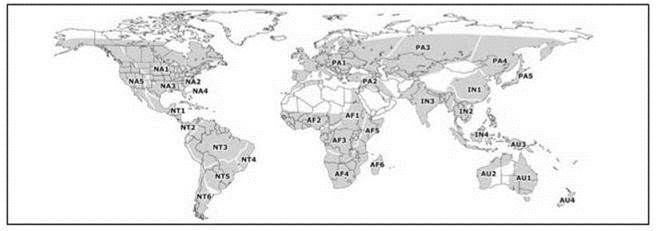
Рис. 4.11. Схема биогеографического районирования пресных вод Graf, Cummings (2007) по распространению наяд.
Области (подобласти): NA — Неарктика (NA1 — Внутренняя, NA2 — Атлантическая, NA3 — Мексиканского залива, NA4 — Флоридская, NA5 — Тихоокеанская); PA — Палеарктика (PA1 — Европейская, PA2 — Ближневосточная, PA3 — Центральноазиатская, PA4 — Амурско-беренгийская, PA5 — Японо-сахалинская); IN — Индотропическая (IN1 — Янцзы-Ганга, IN2 — Индокитайская, IN3 — Индийская, IN4 — Зундо-филиппинская); AF — Афротропическая (AF1 — Нильская, AF2 — Западная, AF3 — Конго, AF4 — Южная, AF5 — Восточная, AF6 — Мадагаскарская) NT — Неотропическая (NT1 — Мезоамериканская, NT2 — Трансандская, NT3 — Амазонки и Ориноко, NT4 — Атлантическая, NT5 — Парано-парагвайская, NT6 — Патагонская) AU — Австралозийская (AU1 — Восточная, AU2 — Западная, AU3 — Новогвинейская, AU4 — Новозеландская).
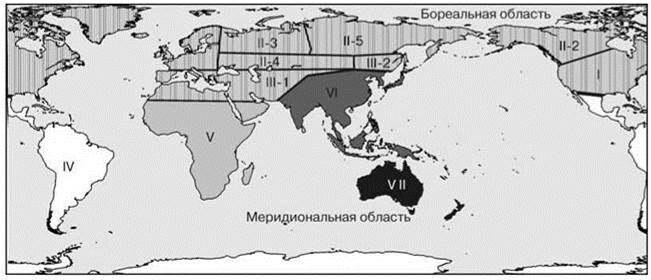
Рис. 4.12. Схема биогеографического районирования пресных вод Белышева, Харитонова (1981) по распространению стрекоз.
Подобласти (провинции): I — Сонорская; II — Голарктическая (1 — Канадская, 2 — Европейская, 3 — Европейско-Сибирская, 4 — Монголо-Казахская, 5 — Сибирская); III — Субголарктическая (1 — Средиземноморская, 2 — Восточно-Азиатская = Амурская); IV — Неотропическая; V — Эфиопская; VI — Ориентальная; VII — Австралийская.
Ситуация осложняется тем, что ценофильные виды — преимущественно облигатно пресноводные и периферические, тогда как ценофобные — преимущественно амфибийные, т. е., как показано выше, обладают различными способностями к расселению.
Биогеография пресных вод строилась в первую очередь на основе изучения рыб. Прочие группы использовали в значительно меньшей степени, прежде всего из- за того, что их таксономия разработана значительно хуже. Многие группы насчитывают гораздо меньшее число видов и/или имеют более ограниченное распространение или наоборот, виды имеют весьма широкое, чуть ли не всесветное распространение, это снижает их ценность для биогеографических построений.
Как уже говорилось, ЦС возникает только на крупных территориях, условия жизни в которых в течение длительного времени достаточно постоянны. Рассмотрим с этой точки зрения пресноводные водоемы. Несмотря на колоссальные площади, которые занимают водосборные бассейны крупных рек (Табл. 4.1), площадь самих пресноводных водоёмов незначительна. Суммарно пресные водоёмы занимают чуть более 1% поверхности суши. Площади морского дна и пелагиали превосходят площадь пресноводных водоёмов ещё более.
Общеизвестны схемы зарастания озера, превращения его в лес. Аналогичные процессы происходят и с речушками и ручьями. С.М. Разумовский (1982, 1999: 529) так описывает процесс исчезновения мелких рек в одном из районов Подмосковья (Рис. 4.13): “в 1880-х годах леса были вырублены почти целиком. В настоящее время она (территория — ИЖ) вся покрыта лесом. Несмотря на хорошо развитый рельеф, эрозионные явления сейчас совершенно отсутствуют, а гидрографическая сеть представлена единственной речкой (3-го порядка по отношению к реке Москва), находящейся на стадии отмирания. Она разбилась на “бочаги”, связь между которыми восстанавливается лишь весной... Однако специальные исследования обнаружили очень развитую погребённую сеть речек и ручьёв, густо покрывающую некогда безлесные склоны... водотоки функционировали, по крайней мере, несколько десятилетий назад”.
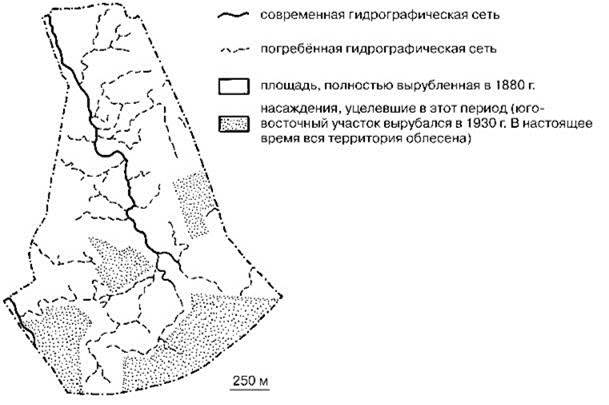
Рис. 4.13. Влияние облесённости на гидрографию в Пионерском лесничестве Московской области (по: Разумовский, 1982).
Таким образом, мелкие реки и озёра являются эфемерными стациями, в которых невозможно образование самостоятельных ЦС. Их можно рассматривать и как пионерные стадии наземных сообществ, и как площадь, населённую ценофобами и пионерными стадиями пресноводных ЦС. Но, учитывая эфемерность и сукцессионную динамику маленьких водотоков и водоёмов, это скорее ранние стадии развития наземных ЦС с примесью пресноводных видов-ценофобов. Поэтому пресноводные ЦС занимают только часть площади пресноводных водоёмов. При этом на значительной части даже постоянно существующих водотоков нарушения столь обычны и постоянны, что сукцессия не может зайти далее нескольких первых стадий. Из-за эфемерности большей части водоёмов формирующиеся в пресных водах ЦС сравнительно с наземными и морскими бентическими обычно гораздо менее зрелые. Именно по этой причине инвазионные виды проникают в ненарушенные пресноводные ЦС гораздо легче, чем в ненарушенные морские и наземные ЦС.
Резюмируя можно сказать, что как ЦС следует рассматривать не отдельные озёра (за исключением некоторых гигантских) или реки, а их совокупность. Аналогично тому, как вид существует в форме отдельных особей, биотоп пресноводных ЦС — отдельные озёра, старицы, реки, ручьи и речушки, однако климакс достижим только в крупных реках и озёрах.
В отличие от наземной и морской среды пресная вода сильно фрагментирована. Пресные воды разбиты на огромное количество в разной степени изолированных участков — водосборных бассейнов отдельных рек, разделённых водоразделами. Её можно представить, как архипелаг, в отличие от непрерывного океана или почти сплошного пространства суши.
Это приводит к нескольким следствиям.
1. В каждом водосборном бассейне аллопатрически могут формироваться свои виды.
2. Бассейны эти неоднородны: условия в низовьях крупных рек существенно отличаются от условий в верховьях.
3. При климатических изменениях возможности для миграции в более низкие или более высокие широты ограничены. Поэтому распространение пресноводной биоты гораздо более связано с историей таксонов, чем наземной.
4. Необходимый рост биоразнообразия при формировании пресноводных ЦС может происходить двумя путями: за счёт видообразования и за счёт вселения в пресные воды морских видов. Последний путь реализуется в районах, где аборигенные пресноводные таксоны отсутствуют или немногочисленны, например на островах. Поскольку морская биота гораздо более разнообразна, чем пресноводная, повышенная доля недавних вселенцев в пресные воды наблюдается также и там, где аборигенная биота попадает в условия, приближающиеся к границам их экологической амплитуды, а миграция из областей, где такие условия нормальны, затруднено или невозможно. Для тропических районов это — умеренные районы, для умеренных — арктические и тропические.
Наблюдаемая в настоящее время гидрографическая сеть в недавнем прошлом (плейстоцене) имела довольно сильно отличающуюся конфигурацию. Весьма часто (в геологическом смысле) происходит перехват верховьев одной реки другой (river capturing). Подобные процессы обычны как в горных системах, так и на обширных равнинах (Валдай, Амазонская низменность и прилегающие территории, верховья Нила и др.), где нет выраженного водораздела. В качестве примера такого изменения можно назвать сравнительно недавнее образование стока Байкала в Енисей, вместо существовавшего ранее в Лену. Другой пример — образование стока верхнего течения р. Конго в нижнюю её часть вместо существовавшего ранее стока в Нил. Однако перехват верховьев способствует обмену сравнительно бедных биот верховьев, тогда как более богатые биоты низовьев в этом процессе, обычно, не участвуют. Их обмен происходит, преимущественно, во время регрессий моря, когда объединяются нижние течения соседних рек. Внутренние моря, являющиеся ныне барьером, ранее бывали пресными и служили путями расселения (Балтийское, Чёрное, Средиземное). В особых условиях опресняться могут и окраинные моря. Так, все северные европейские и сибирские водосборные бассейны, начиная от Енисея на запад до Волги, во время плейстоценовых оледенений неоднократно объединялись системой огромных ледниковых плотинных озёр (Рис. 4.14). При сравнительно небольшой средней глубине (<100 м) площадь этих озёр достигала огромных размеров. Например, озеро в низовьях Оби и Енисея 60—50 тыс. лет назад имело площадь почти в 1 млн. кв. км, т. е. в 30 раз больше Байкала (Mangerud et al., 2004). Периодически эта система имела связь и с водосборным бассейном Понто-Каспия, образуя, вероятно, величайшую из существовавших рек на Земле, длиной превосходившей Нил или Амазонку. Её водосборный бассейн включал всю Западно-Сибирскую низменность, Восточную и часть Западной Европы и реки бассейна Арала, Каспия и Чёрного моря, Центральной и Малой Азии (Mangerud et al., 2004). С другой стороны, даже биота одной реки может сильно различаться из-за наличия крупных водопадов, непроходимых для облигатно пресноводных видов. Например, состав последних в р. Замбези выше и ниже водопада Виктория весьма различнен.
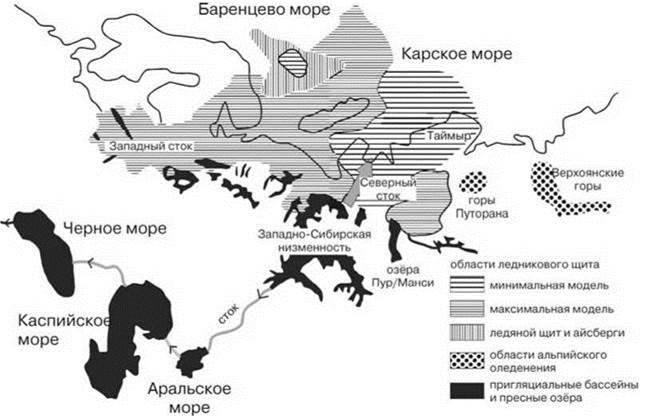
Рис. 4.14. Поздневалдайское оледенение на территории бывшего ЦССР (по: Dawson, 1992 из Аладин, Плотников, 2004).
Другой причиной, объективно усложняющей биогеографическое районирование континентальных водоёмов (понимаемого как районирования ЦС), является наличие эфемерных водоёмов, не связанных с гидрографической сетью. К таким водоёмам относятся лужи, водоёмы в дуплах деревьев, пазухах листьев, кувшинах Nepentes и т. п. Население подобных водоёмов не связано с населением местных ЦС, но разделение их требует досконального знания биологии отдельных видов и при обзорных работах практически невозможно.
В качестве минимального биогеографического выдела обычно рассматривают водосборный бассейн, иногда мелкие водосборные бассейны объединяют. Исключение делают лишь для крупных озёр, которые обычно рассматривают в качестве отдельных выделов. Таким образом предлагаемые схемы районирования построены методом сравнения списков биот и имеют все его недостатки. В частности, биогеографическую однородность этих выделов постулируют, но не доказывают.
Предложенные схемы биотического районирования континентальных водоёмов фаунистические. Причина этого не только в том, что сосудистых пресноводных растений насчитывают всего 2614 видов (2% от видов животных см. Табл. 4.8) в примерно 412 родах (Chambers et al., 2008). Главное — эдификаторная роль макрофитов в континентальных водоёмах значительно меньше, чем в наземных ЦС. К тому же большая часть видов растений отчётливо местного происхождения и отражает сравнительно недавние региональные события и поэтому малоинформативна для глобального биогеографического районирования. В то же время ареал отдельных широко распространённых видов охватывает несколько зон, иногда он почти космополитический (ряска Lemna minor, тростник Phragmites communis).
Доля ценофобных видов в разных таксонах также по-видимому неодинакова. Поэтому схемы районирования, построенные на основе таксонов с преобладанием или существенной долей ценофобов отражают специфику распространения этих таксонов в большей степени, чем специфику ЦС. При биотическом районировании (методом сравнения списков биот) существенно преувеличивается роль ценофобных видов, многие из которых — насекомые). В относительно зрелых ЦС (например, ЦС крупных озёр) роль насекомых существенно меньше, часто незначительно или пренебрежимо мала. Поскольку доля ценофобов выше среди вторичноводных видов, чем у первичноводных, районированию по первичноводным организмам следует придавать больший вес, нежели районированию по вторичноводным.
Из существующих схем районирования континентальных водоёмов наиболее детально проработана схема румынского ихтиолога Petru Banarescu (1990, 1992, 1995), описанная им в 1617-страничной трёхтомной монографии “Zoogeography of fresh waters” (нумерация страниц сквозная). Монография эта исключительно богато иллюстрирована: в ней 401 карта, причём на картах показаны не только ареалы таксонов (до 13 на одной карте), но и часто — их филогенетические отношения. В ней проанализировано распространение рыб, пресноводных моллюсков, высших и некоторых низших ракообразных, ручейников, подёнок, водных клещей и ряда других групп. Эта монография положена в основу данного раздела, и ссылки на неё большей частью опущены. Регионы и субрегионы P. Banarescu в дальнейшем тексте для унификации терминологии названы областями и подобластями. К сожалению, как это обычно в биогеографической литературе, методические вопросы освящены им недостаточно полно, не всегда ясны критерии выделения районов и проведения границ, что делает выделенные регионы и его схему не всегда хорошо обоснованными. К тому же, P. Banarescuкатегорически отвергает возможность симпатрического видообразования, что заставляет его делать сложные и не всегда правдоподобные предположения о развитии биоты того или иного района.
В отечественной литературе широкую известность получили также схемы районирования пресных вод Земли на основе моллюсков Я.И. Старобогатова (1970) и Евразийской части Голарктики на основе рыб Л.С. Берга (1949). Они также учтены при составлении данного обзора. Надо отметить, что биогеографические построения Я.И. Старобогатова сделаны на основе его собственных таксономических представлений, которые мало кто в мире разделяет. Так, автор последнего обзора по биогеографии двустворчатых моллюсков A.E. Bogan прямо пишет, что “we have chosen to ignore... the overinflation of taxonomic levels by the Russian malacologist of the Starobogatov school” (Bogan, 2008: 140). Кроме того, для обоснования рангов биогеографических выделов Я.И. Старобогатов использовал индекс Престона, дающий весьма сомнительные результаты (Жирков, 2001; Азовский, 2010). К тому же моллюски, хотя и составляют существенную часть пресноводных ЦС, не являются единственной группой пресноводных организмов. Анализ географического распространения других таксонов нередко даёт весьма отличную от моллюсков картину. Так, Я.И. Старобогатов выделил в отдельную область озеро Танганьика, в отдельную провинцию озеро Малави, а озеро Виктория объединил в одну провинцию с несколькими соседними озёрами. Между тем число эндемичных видов рыб в озере Виктория в то время, когда Я.И. Старобогатов писал свою докторскую диссертацию (на основе которой и была создана упомянутая монография) было в полтора раза больше, чем в озере Танганьика. Распространение насекомых сильно отличается от распространения облигатно пресноводных рыб, ракообразных и моллюсков. В последнее время предложены схемы по распространению стрекоз (Белышев, Харитонов, 1981, 1983) и ручейников (Moor, Ivanov, 2008). На мой взгляд, общее биогеографическое районирование должно отражать не особенности распространения отдельных групп, а характеризовать биоты в целом, причём предпочтительно отражать распределение ЦС, а не представлять результаты некритического сопоставления сводных фаун, т. е. фаун районов выделенных не по биотическим критериям и гомогенность которых не доказана. В этом отношении схема P. Вanarescu также выглядит предпочтительнее. Поэтому схема Я.И. Старобогатова использована только как дополнительная.
В недавно опубликованной коллективной монографии, созданной в рамках проекта The Freshwater Animal Diversity Assessment (FADA) (Hydrobiologia, 2008) анализ распространения пресноводных таксонов делают путём сравнения биографических регионов, a priori выделенных C.B. Cox (2001). Такой подход следует признать неприемлемым потому, что схема биогеографического районирования должна быть итогом, а не предложена a priori. К тому же схема C.B. Cox основана на анализе распространения наземных млекопитающих — т. е. компонента совершенно иных ЦС. Хотя схемы биогеографического районирования суши и пресных вод во многом сходны, они всё же не идентичны, а в некоторых отношениях различаются кардинально (Табл. 4.8). Тем не менее, несмотря на очевидную неадекватность подхода FADA, этот 637-страничный обзор 58 таксонов (от растений до млекопитающих) широко использован здесь. Некоторые авторы этого обзора, по-видимому также не удовлетворённые прокрустовым ложем схемы C.B. Cox, предложили свои схемы районирования, существенно отличающиеся от схемы C.B. Cox. Наиболее детальна схема C. Leveque с соавторами (2008) по рыбам, схема эта мало чем отличается от схемы P. Banarescu. В то же время опубликованная в этой же монографии схема F.C. de Moor и В.Д. Иванова (Moor, Ivanov, 2008), построенная на основе анализа распространения ручейников (цветная вкладка), весьма сильно от них отличается. Эти схемы также учтены в приведённом ниже обзоре.
Начатый по инициативе WWF (Abell et al., 2008) проект по созданию новой карты биогеографии континентальных водоёмов основан на распространении рыб, большую часть из которых следует считать ценофильными компонентами ЦС. В результате многосторонней оценки распространения рыб, включая не только их ареалы, но и видовой состав, все континентальные водоёмы сгруппированы в 830 первичных выделов. Однако на данной стадии развития проекта никакая группировка этих выделов не произведена. Это определило ограниченную ценность данного проекта для нашего обзора.
P. Banarescu выделяет восемь областей. Из них в Ориентальной, Австралийской, Новозеландской областях и в большей части Карибского переходного региона пресноводная фауна включает много периферических таксонов. Пресноводная биота Индо-Вест-Пацифического региона (как, впрочем, и других островов, не имевших связи с материками) почти исключительно периферическая. Все области, которые принято рассматривать как тропические (Ориентальная, Южноамериканская, Эфиопская, Австралийская), кроме тропических включают субтропические и даже умеренные регионы.
Изученность разных регионов очень различна. Наиболее хорошо изучена Голарктика, в результате во многих таксонах число таксонов, отмеченных только из Голарктики, выше, чем из любого другого региона (Balian et al., 2008b). Нет никаких сомнений, что во многих случаях это — артефакт. Вместе с тем, некоторые таксоны в умеренных широт действительно не менее разнообразны, чем в тропических, а некоторые широко распространённые и разнообразные в умеренных широтах таксоны в тропиках (не в низких широтах!) отсутствуют.
Число ЦС в пресных водах из-за их сильной фрагментированности и сравнительно слабой оформленности оценить сложно. В качестве оценки можно принять число выделенных экорегионов (Abell et al., 2008) — 830. Число пресноводных ЦС, возможно, немного больше, поскольку в крупных озёрах, которые были приняты за один экорегион, ЦС несколько. Впрочем, возможно, что некоторые ЦС разделены на несколько экорегионов. Вероятное число пресноводных ЦС — 500—1000 — как минимум половина ЦС Земли, но структурно они развиты в наименьшей степени.
Обзор биогеографических областей и их подразделений начнём с материков, образовавшихся в результате распада Гондваны, поскольку их биогеография и история проще.